
The Layman’s Guide to
The Amazing but Totally True . .
. Scientific
Facts of Creation
By
Wendy S. Scott
Updated 7/27/09
Contents:
The Fossil Record
A History of
Paleontological Mysticism
Cavemen and Ancient
Civilizations
The
Fossil Record: Brief List of Facts
1)
The fossil record does not substantiate
evolutionary transitions
2)
Fossil record shows species stasis
3)
More variety in past then present (entropy)
4)
Cambrian
explosion—no fossil links
5)
Vertebrate explosion—no fossil links
6)
Tetrapods—no agreed upon fish/amphibian candidates
7)
No ancestor for all three amphibian
body types
8)
Mammals: no
fossil history for development of complex features
9)
Reptiles to mammals—gaps and overlap:
no agreed links, over “100 million year” dormancy.
10)
Fossil
progression interpretation is completely interpretive
11)
Unrelated animal groups inexplicably
share complex traits
12)
The eye is fully formed by first
evolutionary level
13)
Marsupials fossils are widespread with
no history
14)
No fossils to show whale evolution
15)
No Fossil history to show inexplicably
risky transitions
16)
Species do not come up with new genetic
material to survive—they just die
17)
No fossil history for progress of
plants
18)
No fossils to show progress of insects
19)
The dino to bird transition is
backwards
20)
New Link “Ida” sensation too old for a
human link
20)
No link between 3 ½ foot Lucy ape and
man agreed upon by even evolutionists
21)
Hominid
gaps and overlaps contradict Evolutionary predictions
22)
Neanderthals are finally acknowledged
as fully human
23)
Neanderthal DNA tests contradict
supposed age
24)
“Cavemen” and Ancient civilization fit
within the young earth progress
25)
Unchanged “living fossils” defy
evolutionary premise
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Disclaimer: The author of this guide is not a research
scientist. This information has been
compiled from an abundance of easily accessible and confirmed scientific
authorities. The majority of the
information is common knowledge in the scientific realm, while lesser known
facts are cited. Do not quote the author as a scientific authority. This guide is intended to systematically
build the case for Biblical Creation through the logical alignment and
application of the abundance of established scientific facts.
True Blue
All undisputed facts in this guide are in
blue.
The Great Universal Law
“For the earnest
expectation of the creature waits for the manifestation of the sons of God . .
. Because the creature itself also shall be delivered from the bondage of
corruption into the glorious liberty of the children of God. For we know that all of creation groans and
travails in pain together until now . . .even we ourselves groan within
ourselves, waiting for the adoption, meaning the redemption of our body.” Romans 8:19-23
Before continuing with the scientific evidence
that supports Creation, let’s indulge in a discussion of the philosophical
nature. Right here, we are about to
cross the line between creation and destruction. The first two sections address the creation of
the universe, and the Special Creation of life, including humans. We are about to engage in discussions that
focus on the destruction of life and the effects of this destruction on the
earth. Evolution considers creation and
destruction to be two parts of a natural cycle, but the truth is that these
realities are incompatible.
This is the same concept discussed earlier in
the section on Physics. It is
fundamentally impossible for order and precision to increase in the midst of a
system burdened with increasing disorder and disarray. Entropy overrides complexity. Likewise, generative processes that could
create and elaborate on life are thwarted under the dominating oppression of
the simultaneous process of destruction.
When scientists attempt to devise a purely naturalistic, or
self-elaborating explanation for life and the universe as we observe it, they
cannot account for the existence of these two contrary forces.
Simply put, life and death are mutually
exclusive. Destruction does not lead to
the creation of life unless the process has already been set up as a
system. Only the genetic programming
within a seed uses its destruction to produce a tree.
There
is, however, no scientific principle that resolves the contradiction between
the creation of the universe and life within the reign of destruction. The co-existence of life and death defies a
naturalistic origin because destruction always prevails. Everything is destroyed, or dies, but not
everything lives.
Even the remarkable efficiency of the “cycle of
life” and death only explains how the world persists within the present
system. Currently, everything
lives off of the life of everything else, and the death of one organism is
efficiently put to use toward the life of another. The system of death pervades the world,
requiring a well-balanced economy to keep the planet from being over burdened
by it. The usefulness of this system
now, however, cannot explain how it initially came to be, and why life benefits
from death. This efficiency is superior
evidence of design.
We observe the present operations of life, but
we cannot effectively apply these same operations backward, to a time unseen
and undemonstrated, as a process that could have created life and produced the
complexities we presently observe. Death
does not explain life. How something
dies gives no indication of how it was born, yet destruction (from chaos, to
mutation, to death) is evolution’s catalyst for creation. Our observations of the cycle of life can
only enlighten us on this conservation principle, but not on how life began in
spite of death. How did life prevail
to conceive in the womb of death? Or why
did death enter the primordial paradise?
Think about it.
Why do we die? We are all
going to die. You know that, don’t
you? There is no argument, no
discussion. The body fails for everyone,
and it is inevitable. But how can things
seem so good, and yet the worst thing we can imagine is headed straight for
each one of us? Beauty and rot; life and
death. It is a fact that looms over us
all of our waking days, and yet this fact lurks among us unquestioned.
There must be a truth that somehow encompasses
both of the realities of life and death.
Creation and destruction. Science
can’t offer it. Even the faiths of the
world cannot seem to put it together.
Although all recognize the fact of death, they do not address the cause
of its existence. Only the Bible
adequately addresses this relationship and explains it to us.
God made the universe and the world, and all
that was in it, and it was all very good.
He made humans as the head and crowning glory of His creation. He blessed them, but told them there was only
one thing that they were not supposed to do, but they did it. This was called sin, which is to oppose God’s
will or nature.
When they sinned, they did the first destructive
thing in Creation. This opened the door
to all sin, and all types of destruction.
Once Creation was corrupted, it was burdened by the curses of
sin—destruction and death. Therefore,
we, and everything in the physical universe, die, or degrade because there is
sin corrupting Creation, which affects everything.
It is not possible to affect only a few
things. Either everything is universally
subject to destruction, or the universe, and everything in it, is immune to
destruction. Everything touches
something and affects it, and everything is subject to the same laws. The Apostle Paul termed it this way—that sin
is like leaven, or yeast, and when you put yeast into dough, it does not
contain itself, but permeates and corrupts the whole lump (I Cor. 5:6-10).
Other faiths recognize death, and other faiths
recognize a type of sin, or wrong behavior, but none of them relate sin
directly as the natural and logical source of death. Death is so utterly realized, that attempting
to explain its universality seems pointless.
Death just is. We tend to
overlook why death exists, for the more poignant question of what comes after
death. By the same token, we see
sin, or the concepts of impurity as something we must control in order to have
a better life now, or to provide for a better existence after we die. The Judeo/Christian explanation unifies these
two desolating forces—sin and death—with the beauty of creation and life. Sin in creation brought death to every
life. Sin in every human heart is the
cause of death in every human.
“The soul that sins, it shall die.” Ezekiel
18:4
The existence of sin in the world imposes death
on all of Creation. Is there a better
explanation? Yet we would never think of
it. We were not looking for it. God, through His revelation to us, explains
the reality of these two consuming forces, and in His sovereignty, He provides
the answer for both of these oppressors through His Son.
“For as by one man’s disobedience many were made
sinners, so by the obedience of one shall many be made righteous.” Romans 5:19
“For as much then as the children are partakers
of flesh and blood, He also took part of the same; that through death he might
destroy him that had the power over death, that is, the devil; and deliver them
who through fear of death were all their lifetime subject to bondage.” Hebrews 2:14
“And you, being dead in your sins. . .has He
quickened (made alive) together with Him, having forgiven you of your
trespasses; blotting out the handwriting of ordinances against us. . .and took
it out of the way, nailing it to His cross.” Colossians 2:13-14.
“For the wages of sin is death, but the Gift of
God is eternal life through Jesus Christ our Lord.” Romans 6:23
If sin is the cause of universal death, then we
can recognize our need to be freed from both.
The beauty of this message draws us into understanding. In its simplicity, we can grasp the
causational relationship between sin and death both philosophically, as well as
scientifically. Further correlation
between the curse of sin and the burden of death was overtly manifest in the
worldwide Flood—God’s condemnation of rampant, unredeemable sin. The resulting fossil record and the
geological features around the world are powerful evidence of the Biblical
paradigm. These two genres, philosophy
and science, both confirm the truth found in the Creationist model.
Only the Bible answers our intellectual
questions about the intrusion of death, which is confirmed through the
scientific evidence of destruction and the pervading law of entropy. Here, where the relationship between sin and
death is revealed, we find a worldwide event of massive death and destruction
incurred by sin. The clear scientific
viability of this account as documented in the geological evidence establishes
the Biblical model as the more comprehensively sound explanation for how
Creation coexists with destruction than the anti-God, establishment scientific
reasoning does.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
The Fossil Record
“Whereby
the world that then was, being overflowed with water, perished.” II Peter 3:6
In this section, blue may indicate the fact of the existence of fossils,
and the level of strata that they were located at, but does not mean that the
era attributed to that layer of strata by evolutionists is factual. There is less blue in this section because
although the statements truly depict the situation within the evolutionist
scheme, the evolutionist position itself is highly questionable. For example: Evolutionists may claim that
mammals are only found after the Cretaceous, while some mammal fossils may
actually have been found “earlier” in the Permian strata. This situation may be true, but that the
Cretaceous and Permian time periods exist is not a fact, and some evolutionists
may even debate over the implication of the findings. According to the Creation model, all strata
are attributed to the one time deposition by the Flood.
(Much of this section was
gleaned form Evolution: the Fossils still Say No! by Duane Gish)
The Story of the Rocks
The earth offers up its incredible testimony of
the fossil record as evidence of the past.
Most people have been led to believe that the fossil record is the nail
in the Creation Scientist’s coffin, but this confusion is caused by imposing an
evolutionary assumption on the fossils that they absolutely do not reveal. The fact that there is a fossil record at all
validates the cataclysmic Flood so integral to the Creationist model, but that
will be discussed later. The chief
witness for the Creation Scientist might as well be Darwin himself, who
acknowledged the lack of evidence for evolution in the fossils.
In “The Imperfection of the Geological Record”
chapter 10 in Darwin’s book Origin of the Species, Darwin capitulates to
this negative fossil evidence at the end of the first, long paragraph:
But just in the proportion as this process of
extermination has acted on an enormous scale, so must the number of
intermediate varieties, which have formerly existed, be truly enormous. Why then is not every geological formation
and every stratum full of such intermediate links? Geology assuredly does not reveal any such
finely-graduated organic chain; and this, perhaps, is the most obvious and
serious objection which can be urged against the theory.
Darwin frankly discusses a wide variety of
objectionable fossil evidence that he plainly is confounded by. His concluding remarks in the section “On the
Absence of Numerous Intermediate Varieties in any Single Formation” further
concede to:
. . .the absence of innumerable transitional
links between the species which lived at the commencement and close of each
formation, pressed hardly on my theory.
At the beginning of the final paragraph in this
chapter, Darwin recaps in general the most serious detriments in the fossil
record as 1) the lack of “fine transitional forms” 2) the sudden appearance of
“several groups of species” 3) “the almost entire absence” of fossils beneath
the Cambrian strata. Darwin repeatedly
affirms the great breaches in transitional fossil evidence in detail throughout
this chapter, and to some degree in the following chapter “On the Geological
Succession of Organic Beings.” There is
no doubt as to the distinctness of species, abruptness of vast appearances of
numerous kinds, and the lack of fossil evidence to demonstrate the necessarily
“enormous intermediate varieties.”
Though Darwin gives numerous theories as to why such evidence did
not exist, including the anticipation of more discoveries to come, he
admitted in chapter 11 under “The Summary of preceding and present Chapters”:
He who rejects this view of the imperfection of
the geological record, will rightly reject the whole theory.
Though he likely intended the comment to benefit
his own view, Darwin poignantly points out in chapter 10 in “On the Sudden
Appearance of Whole Groups of Allied Species”:
. . . negative evidence is worthless.
This is true.
A lack of evidence is no evidence for a theory. At best, it is neutral, at worst it is
detrimental. He pleads as part of the
reason for this lack of evidence at that time:
We continually forget how large the world is,
compared with the area over which our geological formations have been carefully
examined
Well, things have changed a bit since then. David Raup, curator of the Field Museum of
Natural History in Chicago, once summarized the problem (secondary source
quoted from Gish pg 351):
Well, we are now about 120 years after Darwin,
and knowledge of the fossil record has been greatly expanded. . . ironically,
we have even fewer exampled of evolutionary transition than we had in Darwin’s
time. By this I mean that some of the
classic cases of Darwinian change in the fossil record, such as the evolution
of the horse in North America, have had to be discarded or modified as a result
of more detailed information.
(“Conflicts Between Darwin and Paleontology,” Field Museum Bulletin, Jan. 1979 issue)
Now, more than 150 years later, we have found
hundreds of thousands of fossils, and not one effectively demonstrates a
transition. After what would be hundreds
of millions of years of tortured mutations and natural selection, the fossil
record utterly omits any evidence of this chaotic past, and completely affirms
stasis (species stability in body form).
All species appear suddenly in the fossil record, without preceding
primitive forms, and proceed to remain stable and recognizable throughout the
record even until today. Though species
have become extinct, there is no evidence that they shift into another species.
Today, scientists rest their entire case for evolution in
fossils, but this abundant evidence fails to substantiate any part of the
theory. If there were actual fossils
that demonstrated all the millions of transitions between species, these
bizarre composite creatures would fill our museums. There are none. Moreover, the fossils declare three
fundamental truths that even evolutionists recognize: 1)
there is a lack of documentation for these millions of transitions between the
species layer after layer 2) there has been a tremendous diminishment of
variety in species rather than a trend of increased variety and complexity
since the fossils were created, and 3) there is a lack of change in complexity
between the first appearance of species in the fossil record, and their living forms
of today.
Evolutionists obscure the silence in the evidence by filling
it with the products of their fertile imaginations. Whenever confronted with this baffling lack,
they insist that the earth simply failed to record the information they are
looking for—that the record is incomplete.
However, approximately 98% of living
terrestrial species (except for a lower
percentage of birds) has been documented in the
fossil record, which refutes this
defense. Although evolutionists believe
they understand how life progressed from lower forms, there is still not one fossil that even all evolutionists
believe can prove that this process happened, and there should be
thousands.
Stephen Jay Gould, who again holds to a
non-gradualism view of evolution through Punctuated Equilibrium, sums up the
evidence as presented in the fossil record.
He wrote in Natural History: (secondary citation from Johnson pg
50)
The history of most fossil species
includes two features particularly inconsistent with gradualism:
Stasis. Most species exhibit no
directional change during their tenure on earth. They appear in the fossil record looking
pretty much the same as when they disappear; morphological change is usually
limited and directionless.
Sudden appearance. In any
local area, a species does not arise gradually by the steady transformation of
its ancestors; it appears all at once, and fully formed.
There are hundreds of thousands of fossils of
terminal species, but there is not one fossil that substantiates a single
transition. This should baffle evolution
scientists, whether they hold to Gradualism or Punctuated Equilibrium. Why would only the stable form of
every species be preserved, when hundreds of millions of years of transitions
were supposedly required to reach where we are today? No matter where a species appears in the
fossil record, it appears in its final form without any evidence of previously
more primitive, progressive forms.
Although evolution scientists often label a
species as a “primitive” form, the majority of these “primitive” forms can be
found contemporaneous with the “advanced forms,” just as today’s living forms
co-exist with their supposed “primitive ancestor,” and remain separate
species. A shrew might seem less
complex than a bear because of its size, but not only is this animal’s system fully developed and just as
complex, but behold, we still have shrews and bears. No one suggests that they are related species
today, why would bears and shrews have been related in the past? Without transitional forms to demonstrate such major
structural and physiological alterations, the fossil record reflects the same
situation as the living.
This lack of transitional forms causes evolutionists to
induce most of their relationship conclusions not from the fossil record, but
admittedly from living forms and their similarities. The fossil record
is made out to be the evolutionist’s best weapon, but it is actually the best
weapon against evolution, and an honest look will clearly reveal
this. It is so deplete of incredibly
necessary transitions, that it seems impossible that the evolution scientist is
undaunted by the overwhelming lack of evidence.
Despite this unwavering confidence, there are overt and unbridgeable
gaps between such implausibly linked species that the scientists themselves
can’t agree on a vision of how they could have occurred.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
The Cambrian Explosion
One of the most indefensible examples of such a
gap in the fossil record is in what evolutionists call the
“Cambrian Explosion.” The evolution
model, remember, supposes that after the first single-celled organisms evolved,
one of them began to experiment until it became a multi-celled animal. As discussed in the Biology section, this
would be a tricky process, and there should have been a lot of rudimentary
steps before the new organism stumbled upon the form that would work best. We would expect to discover fossil evidence
to reflect this long period of uncertain body forms until perhaps one or two
basic models appeared. From there, it
would make sense that a tedious process of development would gradually lead to
the radiation of this form into new, but similar body types, and thereby
explain how each species came to be.
The fossil record mocks this concept. Instead of revealing the gradual evolution of
species from one organism, it tells us the story of an abrupt revelation of
species.
Immediately above the strata credited to simple-celled organisms (called
the Pre-Cambrian), lays an inexplicable fossil layer teeming with highly
complex life. There is an absolute lack
of evidence to show transitions between that “first” single-celled creature
(whatever it was—bacteria, algae, or amoeba) and the sudden burst of complex
multi-celled animals. This vast array of
approximately 300 new body types for 35 separate and distinctly recognized phylum,
appears without an inkling in the lower mud of what mysterious creatures they
sprang from.
Some examples of these complex invertebrates are
sponges, starfish, snails, mollusks (clams and muscles), trilobites, sea
urchins, jellyfish, annelids (worms) and great
nautiloids. Nautiloids are one of the
squids in the Cambrian cephalopod family which had a shell that could grow over
9 feet long (quite a remarkable leap from single-celled organisms).
According to the fossil record, the astonishing
squid was actually quite large from the very beginning. Moreover, not only did this squid immediately
perfect the wondrous eye,
but it also discovered the remarkable ability to change color instantly, use jet propulsion,
squirt ink as a defense mechanism (against who knows what since they would have
been the biggest predators at that time in evolution) along with many other
complex abilities unique to just cephalopods.
And they apparently evolved all these advanced gadgets on the first try.
More unfathomable than all this is that the
“upper” Cambrian also revealed an armored fish called the heterostracan, which
is a vertebrate animal with a spine. This is evolution’s first go at
it. How on earth did those little
microbes innovate nearly every body type in one
mysterious leap? All the “Cambrian”
species, which are so divergent from each other, disclose no logical paths from
ancestors to their emergence. They appear together
all at once, nullifying any possibility that
they share an evolutionary interrelationship.
Evolutionists are admittedly baffled by it. One comment in The
Nature Company Guides Rocks and Fossils about a famous Cambrian fossil
site, expresses on page 243 the confusion such evidence creates for
evolutionists:
In
fact, there are more basic types of organisms in the Burgess Shale Fauna than
there are in the world today, a feature that challenges much of our
understanding of the development of life.
Traditionally we have accepted that life gradually gets more complex and
diversified through time, but the Burgess Shale Fauna teaches us that
development of life has occurred as a selection process, with only a few types
of organisms surviving from earlier faunas that were much more diverse. Although the Burgess Shale Fauna is the most
widely known of the Cambrian soft bodied faunas, there are other such faunas
recognized from around the world. . .
Clearly, the fossil record demonstrates there was once a
much greater variety of viable species in the past that we no longer have
today. This confirms that the Law of
Entropy has prevailed throughout biological history, and that species are
narrowing and being lost, not expanded, improved, or gained. How can scientists continue to tout the
processes of evolution when the evidence of their own study causes them to
conclude that this supposed system of gradual biological expansion
managed the impossible, and started at the very beginning with diversity
and diminished from there? How could a
process that necessarily starts with one organism and gradually radiated over
hundreds of millions of years account for the sudden appearance of abundant,
unrelated diversity?
It is impossible for evolution to begin
with an abundant variety, and such a notion contradicts the logical process of
the theory, or any naturalistic explanation.
The evolutionists’ own
assessment reveals that the theory of macro-evolution is contrary to the fossil
evidence. Scientists simply disregard the obvious
testimony of the fossils, and refuse to engage in any discussions of how this
great diversity was invoked without predecessors or experimental history as an
evolutionary mechanism.
A typical textbook that depicts the branches of
evolution will substitute mere dotted lines in the place of actual transitional
pathways. By doing so, they imply that
the transitions are either actual or incidental, when there are actually no
possible ancestral lines to relate these completely new creatures through any
specific single celled organism.
It should be quite insightful that nothing is
found to stand between these “simple” single-celled organisms, and the dozens
of new complex phyla, but little lines.
The very fossil record that should explain our biological history
reveals no history, and from the very beginning of the
evolutionary tale, the evidence demonstrates that these animals have always
been what they are today. In fact, the
“Cambrian Explosion” presented over 1,000 complete species suddenly, while only a small
percentage of them still remain today. This is
not evidence for slow, gradual, upward movement, but for the death of fully
established, complex species and the process of extinction.
More puzzling is that this incredible outburst
in diversity is attributed to the earliest rock strata, but the surviving species
today have remained unchanged in supposedly the “500 million years” since
then. Some of these examples (as
referred to earlier) are segmented worms, snails, clams, starfish, sea urchins,
jellyfish, and the squid (nautiloid). The
fact of this sudden appearance of the “Cambrian system,” and the ensuing
stasis, is direct, unmitigated evidence for the Creation model, and
demonstrates the absence of pressure to affect evolution.
Additionally, the complexity of this abundant
life at this initial stage is astonishing, despite having no
recorded predecessors. An examination of
just the eyes of trilobites and
nautiloids in this earliest appearance of complex forms, unarguably refutes the
millions of years it would have required for them to evolve through mutation
and gradual adaptation. Scientists just
now are beginning to understand the complexity of the trilobite eye from its
fossil remains. The squid eye (or
nautiloid), as mentioned in the earlier section, is comparable to a
human’s—here, at the first burst of evolution!
Without any experimental eye structures or
body types, evolution would have perfected it on the first try, and not a
moment was wasted. There simply is no indication that evolution from
single-celled organisms to these incredibly complex creatures ever happened.
Richard Dawkins, an evolutionary biologist,
confirms this in his book The Blind Watchmaker, pg 229:
. . .
the Cambrian rock strata. . . are the oldest in which we find most of the major
invertebrate groups. And we find many of
them already in an advanced state of evolution, the very first time they
appear. It is as though they were just
planted there, without any evolutionary history.
All of these creatures just appear, worldwide,
fully developed, so much so that the living forms that remain are virtually
undistinguishable from their fossil forms. This
first burst ought to come from millions of years of trial and error, with long
periods of refinement for each species, leaving a remarkable trail of
development. But here, when the workings
of the evolutionary process should be most evident, it is the most silent—and
this is just the beginning. This
well-recognized fact is not evidence for evolution. It is clear, untainted evidence for
Creation. The Creation scientist does not
need to explain the vast abundance found in the fossil record since God created
every kind at once, and filled the earth with an
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Vertebrates and Tetrapods
Following the “Cambrian Explosion”, these
soft-bodied and hard shelled invertebrates suddenly evolved into a vast array
of vertebrate fish. Without the benefit
of the fossil record to demonstrate how creatures with their bones on the
outside, brought their bones to the inside in such a complex structure, we
cannot imagine the physiological transitional functions. There were simply abundant bottom dwelling
(except for the nautiloid) sedentary invertebrate creatures, and then there
were suddenly mobile, completely physiologically different vertebrate
creatures. There is no fossil
indication, whatsoever, of how invertebrates transformed into vertebrate fish.
There is not even a hint of which creature was
the ancestor to the vertebrate fish. Was
it the clam, or the jellyfish, or the sponge, or the starfish, or the sea
urchin, or the nautiloid (squid) or the trilobite? Who can imagine such a thing or what possible
creature gave rise to them? Had there
been some of these intermediate forms preserved in the mud that they were
living in, even relatively few compared to the abundant terminal species that were
preserved, then scientists would not have to imagine them. But since they do have to imagine
them, there seems little reason to accept the transitions as fact. Since the vertebrate heterostracan fish is located in the
upper Cambrian level, right along with the invertebrate creatures, there is no room for the
millions of years of possible transition into this physiologically unique body
type.
Most of the fish we are familiar with today are
established early in the fossil record along with many fully formed,
extraordinary extinct species. Many of
our living species have much larger fossil counterparts, although one would
logically assume that a species’ size would increase with its success, not the
other way around. There is really
nothing more to say about the evolution of vertebrate fish. There simply is no transitional species, real or
imagined, between invertebrates and fish. That’s
it. It is that simple. There are millions of fish preserved in the
strata, and millions of Cambrian invertebrates, but not a hint of how they are
connected
evolutionarily speaking.
It shouldn’t be necessary to point out how
incredibly complex
an animal with an internal skeletal structure is, with the muscles, nerves and
soft tissues on the outside. The spine
alone is a marvel of ingenuity, and to run the sensitive central nervous system
right down through these delicately connected bones, is pure genius. Additionally, there are numerous features of
the fish that perfectly suit it for its unique and mobile life in the
water. Imagine that it would have taken
thousands of steps to develop the new mouth and teeth, plus the special bones, fins, muscles,
swim bladder, a lateral line sensory system, and floating stones in the ears
for balance. Such an incredible transition is completely
mysterious. That there is no trace or
hint of how it happened is a great hindrance even to the imagination. Really.
Go ahead, try it . . .
See?
Obviously, whatever rapid and fortuitous mutations managed to suck that
skeletal structure inside, was too much to bear for the rest of the
invertebrates, and they opted to stick with the old form. The poor creature that evolutionists envision
must have become the first fish apparently lived its life in secret. Since the first two major evolutionary
developments (the Cambrian explosion and the appearance of vertebrates) are completely undocumented,
(and evolutionists do not argue about
this fact) then there is no need to examine the rest
of their claims. Evolution clearly did
not happen because it irrefutably didn’t happen at the first two levels. Not only is there no evidence to indicate
evolution, the evidence indisputably testifies against evolution. Scientists cannot call evolution a fact if
they have to skip past the foundational stages to a better opportunity. But as good sports, let’s continue.
Once fish were fully established,
they were compelled by unknown forces to venture from their well suited
environment, into the world of air and land.
An intriguing mystery for evolutionists should be why a fish would want
to leave the water at that time since according to their own timeline, there
was nothing out there yet to get. No
bugs, no plants. This adventurous fish
would have spent all that energy evolving just to set out and wander the land
out of pure curiosity, hoping to run into some more water with food in it
before it dried out. And of course, it
would have to sense millions of years in advanced that the food was about to
run out in its pond in order to enact the necessary changes in time for it to
venture out and meet its impending needs.
Can you see it sitting there on the water’s edge, dreaming for millions
of years about how it would leave its little pool one day? Perhaps it used mind control to influence its
genetic makeup every generation just for the chance to venture out of its
little mudpool.
This transition from fish to tetrapod
(four–legged animal) is (surprise) wholly undocumented by the fossil
record. Though there are millions of
fish fossils, and thousands of amphibious tetrapod fossils, there are no agreed upon
fossils to help evolution scientists imagine how the transition was
accomplished. Without this evidence, any logical deductions
as to the forms and functions of what would be an amalgamation of the two
species, is illusive and contradictory to the known workings of biological
functions. The remarkable uniqueness of
each species contradicts all imagined evolutionary applications, even if
one does not require actual evidence.
The difference between a water breathing fish
and an air breathing land animal is more immense than that between a gas engine
and an electric engine. Even if they
serve the same purpose, the workings are so incompatible that one would not
consider pouring gas into the electric engine expecting it to function. The entire machine is built around the use of
its energy source. Randomly changing a
single part will not allow it to function.
Adding a gas tank to an electrically powered engine is not sufficient
without fuel lines, cylinders, pistons, exhaust systems, as well as their
affiliated small parts (distributor, spark plugs, intake valve, parts for
regulating the fuel-air mixture, and all the tiny parts that make each of them
up).
This transition is so complex, that one would
hardly undertake to start with an electric engine to transition to a gasoline
engine. We now have hybrids which must
be built from scratch in order to incorporate the two systems. One is not built from the other. If one were to undertake such a thing
though, even the simplicity of this nonliving machine cannot undergo such a
transition while it remains running. This is exactly what scientists suppose the
transition from fish to tetrapod undertook, while remaining functional. Not only functional, but each of these
transitional forms were supposedly better than the initial form, justifying the
evolution. Since we don’t have these fossils for
study, we don’t know how each of these transitions
would have been an advantage.
Scientists have tried to imagine the transition
from gills that already are functional, to air-filled lungs that are also
successful, without a period in between when neither is
successful. Most amphibians undergo such
transformations from tadpoles, but their genetic coding has programmed their
respiratory system for this transition. We also have been
pre-programmed to make this same transition, from receiving our oxygen through
the umbilical cord, to receiving it through the lungs. The tadpole is merely an external embryonic
stage for the frog, and therefore all of its unique qualities are based on the
frog’s genetic programming.
Unlike both fish and reptiles, amphibians
uniquely have smooth skin, and no scales. This in
itself is remarkable since scales and smooth skin are very different structurally, and the amphibian (no scales) is supposed to stand in
the evolutionary gap between fish (with scales) and reptiles (with scales), creating two unnecessary but complex
innovations.
Moreover, amphibians’ unique skin
absorbs and releases oxygen, enabling the preprogrammed transition from gills
to lungs.
This pre-programming, of course, could not be the case in the transition
from fish to Amphibian unless there was an evolutionary plan.
None of the potential fossil (or living)
candidates displays any telling indications of such a transition. Though there exists the lungfish, it also
presents several difficulties today. The only lungfish that has
fins sturdy enough to resemble the start of limbs would be related to the
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
There are three major species of fish currently
debated among evolutionists as the possible ancestor of the amphibian
tetrapod. The reason for the debate is
not that there is such a good selection of candidates, but that there is no
sign of the millions of transitional forms that would have been necessary between
an animal suited for living and swimming in the water, and an animal suited for
living and walking on land. There simply
is nothing to demonstrate that it ever happened. Lungfish are always lungfish and amphibians are
always amphibians in the fossil record, just as they are today.
The three main candidates for ancestor to the
amphibian are the noted lungfish, rhipidistian, and coelacanth, all of which
are fully fish, giving no indication as to how they became an amphibian. The debate over the amphibian ancestor rages
on in light of the lack of clear, certain evidence. Evolutionists acknowledge this lack of
evidence, conceding that there may never be an end to the debate. One specialist in the area, Malcolm Browne,
comments in his March 16th 1993
No
one can be certain which group or groups of fishes was the first to make the
transition to land, or what their evolutionary pathways may have been . . . the
transition from water to land occurred long ago, and various family trees
suggested by the fossil record are so tangled that scientists acknowledge they
may never know definitively.
How is it that scientists continue to call
evolution a fact when they admit to having no evidence, and place the blame for
this lack on how long ago it happened?
Once again, evolution’s lack of evidence is somehow contrived as
evidence in its favor. The fossil record
should have been a little more cooperative in preserving the transitions, as it
inexplicably preserved
all the diversity prior to the supposed transition and
immediately after the transition to amphibian, in abundance. But even if one does not require evidence at
this crucial transition from water to land, there are other issues to
consider.
In addition to the difficulties in imagining the
transitions in respiratory systems, there are numerous drastic structural
innovations necessary to prepare a fish for land. Remarkably, although one might imagine
how fins might become limbs in spite of the fossil evidence, fish are physiologically
incapable of fully supporting their weight on land using their fins. The pectoral fins of fish, like the walking
catfish, are rays of thin bones that project out and anchor the body as it
wriggles. It does not walk. Fish fins do not work as limbs because they
do not have a skeletal bone structure like the feet of tetrapods. There is no structure that resembles limb
bones, nor attachment structures to the skeleton for weight bearing like a
thoracic girdle or pelvis.
Tetrapod limbs are strongly attached to the
pelvis and the spine. The small fin
bones in the majority of fish species are not attached to the spine at all, but
the front fins are generally attached to the skull (imagine if your legs were
attached to your head?) without a suitable, weight bearing attachment or limb
structure. The rear fins are typically embedded in muscle tissue alone, and
receive no skeletal support. Fish have thin, light,
flexible bones, but amphibians and other tetrapods have heavy, strong skeletal
structure to support ever aspect of the limbs and articulate them to the robust
spine.
Ichthyostegid is regarded as the likely first
full amphibian, which is now extinct.
The fish “ancestor” that most evolutionists are convinced best resembles
this first amphibian is the rhipidistian.
This is a type of crossopterygian (such as the coelacanth, one of the
lobe finned fishes). Neither the
rhipidistian nor the coelacanth exhibit any physiological or skeletal move
toward land dwelling. In fact, the unusual coelacanth has been found still living in the oceans
off the coast of
The coelacanth must rationally be removed from
amphibian ancestry since soft tissue studies of the actual fish prove that
there are no physiological transitions occurring in the living form toward
amphibian features such as lungs, circulation, and heart. Most damningly, since these fish have been
studied in their environment, scientists have discovered that their very meaty fins are not used
for walking in any manner. They use them simply as
fins, but in a uniquely very fluid sequence that keeps the body balanced in one
place.
Moreover, coelacanth lives so deep and in such a
bizarre little world, its lifestyle clearly did not lead to a land walker. For good measure, to further spoil the
association, the coelacanth is ovovipaous, or live bearing rather than egg
bearing—very advanced for such an ancient fish considering mammals haven’t
invented it yet. We can see how easy it
is for scientists to impose their own ideas when interpreting lifeless bones.
Most evolutionists believe that the rhipidistian
is the best candidate for the transitional ancestor to amphibians—probably
because at least is hasn’t shown up embarrassingly alive. Since the coelacanth has always been
considered closely related to the rhipidistian, both may be reasonably discounted
as lacking the physiological features (and likely behavioral as well) that
could demonstrate such a transition.
While this fish seems to share a similar cranial structure with
amphibians, the
few bones in these projecting fins do not distract from its otherwise fully
aquatic form. Although the fins are
thicker, and not rayed, like other fish, they completely lack any limb
structure whatsoever, with no bones analogous to the amphibian tibia, femur,
knee joint, or even foot.
The pectoral fins offer the most robust set,
though they are grouped together no better than a clump. None of the potential limbs attach to a pelvis or
thoracic structure to prepare for weight bearing, and the species is completely
lacking the robust ribs of its nearest supposed kin. If this fish is supposed to lead to
amphibians, it has a long way to go.
Like, all the way. A visual
comparison of the skeletons makes this evident.
In fact, none of the fish on
record remotely possess any kind of preliminary limb structure at all, let
alone weight bearing on land.
Recently, a new discovery has breached onto the
public mind like a tsunami wave, called Tiktaalik. This new transition, found in the Canadian
Arctic, purports to be the next step to land dwelling, but all it all, it is
quite fishy. The unusual characteristics
stem from essentially sturdier (though quite short) ribs, internal gills
(rather than external with bony arches) a crocodile-like head and lobed
fins. These lobed fins get the most
attention because they contain several bones, some of which are longer than
others. Discoverers believe that these
are the beginnings of digits and a wrist structure in a move to walk on land.
Just like the coelacanth, these meaty fins do not
actually align into digits, and they are imbedded in the flesh of the fin, not
articulated into limb structures, therefore they are not capable of supporting
the fish on land. Although the shoulder
structure is more pronounced than most fish (though lobe finned fish generally
have this structure), these limbs still do not articulate with the spine, which is necessary for
weight bearing. Additionally, the rayed
fins prevent significant weight bearing as well, and are evidence that the fish
spent the majority of its time swimming.
Moreover, there is a complete lack of development of any
hind limbs,
absolutely imperative in a legitimate transitional candidate. There are only small rayed pelvic fins, with no
legs, hips or any analogous bone structure.
Tiktaalik would then be the opposite of what would be expected of a near
transition to land. On land almost all tetrapod
hind limbs are the strong appendages, while the forelimbs are generally weaker
because the forelimbs do not propel the animal forward—the hind limbs do. It was once believed that coelacanth used its
meaty pectoral fins to walk around in shallow water, but now we know that they
never use their fins in this way. The
evidence offered by Tiktaalik adds nothing structurally to show that it was any
more ambitious.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Some evolutionists have even commented that
Tiktaalik has too
many vertebrae, and they are poorly ossified.
Poorly ossified bones lack the rigidity and strength for land-dwelling
because they are very flexible, and more cartilage-like. Babies must go through this transition of
ossification before they are born, or they could not bear their own weight
outside the womb. Additionally, this fish
was placed in the mid to late Devonian, which is just moments before the first
amphibian (Ichthyostega) appeared—a close call considering how far it has to
go. The very heavy, bony scales(just like coelacanth)
that cover the fish also tell us that it had to stay in the water, and was not in the
process of evolving amphibian skin capable of retaining moisture.
Most significantly, there remain the same gaps
in demonstrating the most significant tetrapod transitions. This fish fails to supercede other lobe
finned fish in manifesting actual advances in limb structure, front and hind,
and adaptations to aid in leaving the water even minimally. Whatever interesting things Tiktaalik was doing,
it was not walking, and it was not leaving the water. Looking a lot more like a coelacanth than an
amphibian, this fish too may turn up one day, swimming its strange little way,
and showing us what we could only guess.
Real life is funny like that.
While none of the fossils offered by evolutionists
solve their major transitional dilemmas, there are some interesting living
animals that give scientists a hard time for classification. Some have been classified as amphibians, but
closely resemble lungfish. These
species, such as the Amphiuma and the Grater siren, are quite eel-like, and
dwell primarily underwater. If these species somehow
fit into the fossil record, and managed to clear up a few mysteries, how nice
that would be for evolutionists.
However, we are not looking for shared traits
among different classes of animals; we are looking for a direct lineage between
a fish without limbs to amphibians with limbs.
We don’t get to take random living species and align them in a
convenient lineage. Since no such
creature is provided by the fossil record, the transition is impossible to
substantiate. Additionally, the lack of
any candidate exhibiting progress toward weight-bearing limbs is further
complicated by the wide range of amphibian body types one fish would have to
radiate into.
Many obstacles arise when attempting to account
for all the different forms of amphibians through this one link. The first problem comes when we note that all of these species of fish
and the supposed first amphibian, ichthystegid, have the arch type vertebrae,
while all other amphibians have the very different spool type vertebrae
structure, making them distinct species without logical link to the “first”
amphibian. It is difficult to imagine how and why the
transition was made to this most vital structure, without inviting certain
peril.
In fact all amphibians are so structurally and
sometimes physiologically different even from each other, that there is little
to inter-relate them, let alone to a common ancestor. What possible intermediate creature gave rise
to the whole amphibian class
composed of species that grew legs (salamanders, and newts), and a species that
lost its legs (caecilians), or a species that reconfigured its legs and made
nearly its whole body look like a giant mouth (frogs and toads)?
The few features amphibians do share, in light
of the difficulties with a common ancestor, could just as easily be categorized
as what evolutionists call “parallel evolution”. This is a concept used to explain the development
of similar features in closely related species when there is no plausible
common ancestor—a remarkable, and unrealistic feat. Although it is more sensible to address all
amphibians through a single fish ancestor, it is pragmatically unfeasible, and
unrealized in the fossil record.
Amphibians, in their wide range of stable forms, simply appear without a
trace, leaving serious questions about this transition from water to land. Evolutionist Robert L. Carroll testifies to
this in his book Vertebrate Paleontology and Evolution, p 180-181:
(secondary citation from Gish, 93)
When they first appear in the fossil record,
both frogs and salamanders appear essentially modern in their skeletal anatomy.
. . Despite their similarities, frogs, salamanders, and caecilians are very
different from one another in skeletal structure and ways of life, both now,
and throughout their known fossil record. . . we have found no fossil evidence
of any possible antecedents that possessed the specialized features common to
all three. . .
Since there are no fossils to demonstrate any of
the thousands of transitional steps between the water breathing and dwelling
fish to an air-breathing, land walking amphibian, then there is no evidence in
the fossil record to support tetrapod evolution at all. This most significant change in the direction
of life, leading to all land animals, offers not a sliver of evidence that it
ever happened. The concept of fish to
tetrapod evolution is based on conclusions drawn from similarities in living
forms, not actual fossils.
The fossils do not tell this story.
That is why the evolutionists are still looking for the puzzle pieces to
make the picture look the way they envision it.
If these evolutionary assumptions are based on
what we observe today, and the fossil puzzle pieces are not there, then
there is no proof because we know as a fact that today’s living forms are
separate and distinct. If there is no
discernable link between fish and tetrapods, then the evolutionary ladder is
stopped again at the next wrung, because all land dwelling, air-breathing
animals, including birds and whales, had to come after this crucial
moment. If there is no undisputed
evidence that it actually happened, then is it logical to say that evolution is
a fact and Creation is a fantasy? Which
one is borne out by the evidence?
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Warm and Fuzzy—Mammals
There are innumerable questions about all
transitions from one species to another.
Although there are many mysteries about the next supposed transition
from amphibians to reptiles, and reptiles to dinosaurs, of great interest to us
is the transition from reptiles to mammals.
Placed by evolutionists some time deep into the age of dinosaurs,
“primitive, true” mammals supposedly first appeared in the mid Jurassic. Evolutionists contend that millions of years
before that, some Permian age reptiles were beginning the long journey of their
metamorphosis into mammals.
There are dozens of
necessary transitions that would be required in order for a reptile to be altered into
a mammal. First, the skeletal features are obvious. The reptile has a sprawling stance, with the
legs out from the body, while mammals have their legs beneath them, (also like
most dinosaurs). This feature makes the
mammal faster, more agile, and gives them a higher endurance. Such a change requires major restructuring to
the pelvis, limb construction, joint angles, and the scapula.
There would also need to be an alteration from
the one-boned reptilian ear to a three boned mammalian ear, differentiation in
the teeth, a change in the number of jaw bones, and the addition of a palate in
the mouth. There would have to be a
change in the number of neck bones to 7, the formation of the mammalian
thoracic girdle, and the compartmentalization of the chest (mammals’ ribs are
joined by a breastbone, reptiles’ ribs are not). These few adjustments are just the
start.
There are even more internal, or physiological
alterations that would be necessary as well.
Most of these soft tissue features are not detectable in fossilized
bones, so any such transitions would be covert.
Examples of these necessary
transitions are shifts from scales to skin with accompanying sweat glands,
nerves, blood vessels, and fur. The
creature would need to produce a diaphragm, completely renovate the
reproductive system from egg laying to live birth, and invoke mammary
glands.
The most drastic change, however, would be from a
cold-blooded to a warm blooded metabolism.
This complex innovation involves the entire body system of nerves,
circulation, blood pressure, digestion, fat storage, heat regulation and
generation features, and many more factors.
There would be physiological adjustments to the kidney, the structure of
the heart, and a general increase in the muscle’s capacity to do work. The entire body would work under a completely
new system, as nearly innumerable alterations would be required.
Such a feat is not only overwhelming, but
incomprehensible. A reasonable question
might be how the genetics managed to utterly renovate an entire body’s systems
into a completely new direction, toward one perfectly balanced goal, by total
chance. The next reasonable question
should be why? Evolutionists agree that such drastic and multiple physiological
changes would have to be gradual.
Moreover, individually, each alteration would be gratuitous without all
the other elements already in cooperation.
Since such a transition is unimaginable, in order to regard it as a
fact, some reasonable substantiation should be required.
Since the physiological transitions themselves
can’t be tracked, the only way to infer if these changes could have happened is
through fossil finds that painstakingly align to demonstrate the major skeletal
transformations through a traceable lineage.
If the fossil skeletons do not clearly demonstrate a lineage of this
long gradual shift, then logically there could not be an opportunity for the
more complex transformations physiologically.
Do the fossils provide this substantiation?
Evolutionists propose that mammals may have
developed from a loosely associated group of animals they have labeled
“mammal-like reptiles.” Members of this
group display one or more of the important mammalian skeletal characteristics
that paleontologists look for, chiefly in the skull features, or tooth and
palate structure. Evolutionists believe
they detect a more upright walking stance in the angle of some mammal-like
reptile joint angles verses the wider, squat reptilian stance. This feature, as well as the others, suggests
to them that mammal-like reptiles were on their way to becoming mammals.
The other skeletal earmark features that
paleontologists typically look for to demonstrate this transition are missing
in mammal-like reptiles. There is no
indication in these
candidates a of transition from multiple jaw bones in reptiles to a single
jawbone in mammals, and a single ear bone in reptiles to three ear bones in
mammals, which would be vital for validating reptile to mammal evolution.
Evolutionists must believe that mammals evolved
from reptiles because there would have been nothing else for them to evolve
from. Most of the specimens placed in
this wide category of mammal-like reptiles, however, do not skeletally resemble
mammals in the slightest. There are some
strange creatures associated with the group, like the Dimetrodon, which is a very odd reptilian
specimen known for its high sail of extended spines. The
fossils placed in the mammal-like reptile category are so widely varied, that
there is no relationship evident among them even to each other, let alone to
any specific mammals.
Candidates are frequently placed in this group
because of one of the features mentioned, even when the rest of the creature is
very un-mammal like. They are
essentially categorized based on the number and location of openings in the
skull, and tooth structure. The vast
variety of body types within the mammal-like reptiles ranges from dog size to
hippo, from predator to grazer, making a line of ascent completely inferred
across improbable gaps. The illusion of
progress is created by the group, and not by the characteristics of individuals
since its members may each exhibit one desired characteristic, but not all, and
they cannot be aligned in a feasible progression.
Most mammal-like reptiles have obvious features
that completely disqualify them from even a superficial link to mammals in
general. All but a few in this category retain the
reptilian ribs that extend nearly the full length of the body, without the
compartmentalized rib cage necessary to demonstrate the development of a
diaphragm. Any candidate that does not
have a diaphragm could not be developing a warm-blooded metabolism because it
is vital for the necessary rapid gas exchange.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Major differences, such as size, skull shape, and overall
skeletal configuration have prevented a reasonable connection between any
specific mammal-like reptile to any specific mammal. Though one or another mammal-like reptile
might have shared characteristics with one or another mammal, other features,
including recessive reptilian features, like teeth or jaw bones and joints, eliminate a feasible evolutionary line to any “early”
mammal. In reality, much of the skeletal
structure and stance seen as mammal-like, can also resemble dinosaurs, blurring
the line of ascent even more.
Paleontologists have narrowed down the
mammal-like reptiles to a subgroup of therapsids called cynodonts because of
their teeth differentiation. Mammals
have different sized teeth for different functions, such as incisors, molars,
canines. Reptiles generally do not, so
an advancement in this feature is necessary to move toward mammals. This type of shift seems incidental compared
to the major physiological shift from reptiles to mammals, but necessary.
Most cynodonts in this category are about the
size of a large dog. Two of the
candidates, however, are the Thrinaxodon, which is almost 2 feet long, and
cynognathus, which is just over 3 feet long. Although
the skull, teeth and rib configuration in cynodonts may be closer to mammals
than all the others, these species still have a reptilian jaw and jaw joint. Additionally, their teeth do not meet in occlusion
like mammals. The recessive reptilian
features, and a lack of predecessor or descendent pathway isolates these
unusual animals in their own category.
Overall, the cynodonts are too large to be even
a near link to what evolutionists conceptualize as the first true mammal. The shrew is the first, and primary mammal
believed to exist for nearly 160 million years until the Cretaceous. The shrew that is credited with this first great
leap to mammalhood, is very small, with a distinctly sleek body, and only about
4 inches long. The immense gap between the most mammal-like
reptile, and this first “true” mammal is too vast, in both size and structure
with an overall lack resemblance between them.
There is no feasible link between reptiles and mammals.
The structural similarities between some
mammal-like reptiles and mammals may make for an unusual group of creatures,
but the differences among them are so great that they are just another class in
the diversity of the animal kingdom.
These shared traits no more link the reptiles to mammals than wings link
birds to bats. Unless there are specific
intermediate fossils to bridge the large gaps, then there is no evidence that
it happened. Scientists all agree that
the group called mammal-like reptiles are an unusual group that we do not see
today. They demonstrate a wide range of
unique features, just like mammals demonstrate a wide range of similar
features. Some features will always be
shared by other species because tetrapods share a similar basic body design,
created to operate under similar conditions.
The gaps in the transition between reptile and
mammal remain evident in that the cynodont still cannot enlighten us about the
intermediary shift between significant reptilian and mammalian features. Structurally, skeletons only tell us so much
about a creature, and because most land animals share the same basic skeletal
structure, it would not be hard to find many similar traits between the two
groups. Because we cannot put a face and
skin on most of the extinct species, we may never know their true identity (who
would imagine that the big hole in the face of an elephant would be for a trunk
unless we had seen it?).
What these bones will never explain is how a
reptile could physiologically transform into a mammal with all the incredible
differences, and why on earth it would bother.
The vast gaps between reptiles and mammals prevent even our imaginations
from constructing the transitions with plausible models. The fossils do not demonstrate the necessary
gradual stages that altering these soft-tissue physiological features would
require. Such a gradual change over
millions of years would generate numerous transitional species for the fossil
record instead of this sparse allocation typical for the evolutionary
evidence.
There is no semblance of a pathway between them
that demonstrates this tremendous alteration to the entire body. There is no evidence of the shift from a
cold-blooded metabolism to a warm-blooded metabolism, or from egg-laying to
placental birth. There is no remotely
established fossil link between reptiles and mammals, there is no logical
propulsion or means for such a transformation, and there is no evidence that
the evolution of reptiles to mammals is a fact.
Just as with the Cambrian and invertebrate explosions, there is no
feasible evolutionary pathway recorded by the fossils. Which is pretty detrimental in light of the
mammal explosion to come.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
The Mammal Explosion
Lurking in the dark obscurity beneath the feet
of giant dinosaurs, a plot is being hatched by the restless DNA of a nocturnal
little creature. It may take 160 million
years, but some day, mammals will get their chance. Their DNA have been planning for this moment,
storing up great ideas for their takeover, and when that moment comes, the
world won’t know what hit it. Boom! Here come the mammals.
The mystery of the exploding mammals is great. According to
evolutionists, the true unveiling of mammals doesn’t occur until the late
Cretaceous, early Tertiary.
Evolutionists rely on the Dinosaur extinction as the trigger that
catapulted the instantly remarkable mammalian diversity. Mammals are considered “advanced” animals,
and if evolutionists cannot present the illusion that they evolved later than
dinosaurs, then there really is no evidence that evolution is a progress to
higher order. What sense is there in
believing in evolution if mammals and dinosaurs all lived at the same
time? That would mean that every animal
group that ever existed lived at the same time, which negates the rationale for
evolution.
However, the only animal group that can provide
any possible ancestor for mammals—mammal-like reptiles—would have lived much,
much earlier. Papa cynodont and his mammal-like
reptile buddies reportedly dwindle to extinction by the end of the Triassic/
early Jurassic—some say as a gradual impact of the Permian extinction. Though the first mammals supposedly appear at the end of the Triassic, only the sparsest record
of these “early” mammals is allotted to the nearly 160 million years that
precede the dinosaur extinction, and mammalian explosion. This forces evolutionists to impose a
lengthy, non-productive gap on mammalian evolution. The evolution timeline is dictated not by evidence,
but by its own necessary presumptions.
This great dormancy in development strains the
relationship between the early mammal-like reptiles, and the quite late “true”
mammals. The incredible gap between the
demise of these mammal-like reptiles and the sudden ambush of “advanced”
mammals is about two geological ages.
However, this remarkable transition (according to evolutionists) leaves
virtually no traceable mammalian existence for nearly 160 million years, when
the time could have been applied to the industrious evolution that would
come.
Supposedly from this trickle of ancestors,
“advanced” mammals then exploded in abundant diversity without warning,
overstepping any substantiated link to the previously abundant mammal-like
reptiles. Considering evolution is based
on the survival of the fittest, how did the floundering legacy of mammal-like
reptiles manage to instigate the explosion of mammals 160 million years after
their passing, through such a sparse population of “early” mammals—too unfit to
leave but a trace? This transition is
hardly inspirational, and yet the best hope for the theory.
According to evolutionists, this marvelous
revolution happened without leaving any recognizable evidence of these millions
of vastly divergent transitions. There
is no evidence to indicate how there were virtually none, and then suddenly the entire
range of distinct mammalian species of all sizes appeared, without transition,
from bears to bats, around the world.
This incredible explosion produced all 32 orders, nearly 100 families,
of mammals distinctly from their first appearance.
More disturbingly is that the little shrew, which looks just like the
little shrew of today, bears the responsibility for
the source of all this diversity.
Evolutionists give a muddled account of how
mammals struggling to survive, for nearly 160 million years, was able to
innovate the inexplicable body types in obscurity so that they could spring
into their vast diversity the moment the opening presented itself. Placentals, marsupials, monotremes, grazers,
predators, swimmers, flyers, and everything in between, were rapidly generated by
the intuitive, and industrious genetics, just because. The astonishing array of new body types is
inspirational. Most of these unfamiliar
animals did not survive until today, ranging from enormous long-necked rhinos,
to giant rodents, and unusual beaver-like gnawers, non-ungulate herbivores and
dozens more.
This incredible array of species would have
needed a long history of development.
The fossil record, however, contradicts this concept, as all these species appear
suddenly without any developmental preparation between the decline of dinosaurs
and the emergence of all these mammals. Even
evolutionists recognize that this great diversity of mammals suddenly appears
without precedent. That is why they need
to explain it with the extinction of the dinosaurs. The Atlas of Life on Earth comments on
page 237:
The Cenozoic Era has become known informally as
“The age of mammals” due to the explosive diversification of this group as the
dominant large-bodied animals. Mammals
had first appeared in the late Triassic period of the Mesazoic, but there were
few opportunities for them to expand into a world dominated by dinosaurs. . .
So what evolutionists are saying is that mammals
actually evolved before dinosaurs (which should upset our sense of the
evolutionary scheme and “advanced” forms).
Then they barely survived with only a few forms to speak of until some
disaster killed all the dinosaurs. Then,
because dinosaurs went mysteriously extinct, mammals decided to kick their
genetic mutations into overdrive so that they could take over the world. Apparently, in evolution, one’s DNA is
intuitive, and will creatively innovate vastly new body types to purposely
advance into vacant territory. Is that
scientifically logical?
Unfortunately, there is absolutely no record in
the fossils to help clarify this incredible expansion of mammals from the
shrew, over time, to their vastly diverse final forms. Any evolutionary graphic that actually
attempts to depict the evolution of mammals shows a separate branch for almost
every family of mammals, rather than an ascending line of mammals progressing
along the same branch. This confirms the
lack of transitional evidence, and difficulty in relating their evolutionary
pathways. Since the whole array of
mammals are placed within the same relative timeframe, assigning progressive
lineage relationships among them is arbitrary.
When one looks at the evolutionists timeline, the age of the
dinosaur up to the mid-Cretaceous is depicted directly before, and intermingled
with this great eruption of mammals in their terminal (fully formed) species in the
late Cretaceous early Tertiary. Both
time and fossil evidence do not allow for the rapid evolution of these diverse
species of mammals to be instigated as a direct product of the decline of the
dinosaurs.
However, according to paleontologists’ own
interpretation of several finds, dinosaurs and a surprising diversity of
mammals existed in the same strata, which would make them contemporaneous in
the late Cretaceous, entering into the Tertiary mammal explosion. This shrinks the available window of
development to nothing in order for the demise of dinosaurs to incite the
explosion of all forms of mammals (from shrews into bears and bats and pigs and
ungulates and hippos, and anteaters, whales and so on). Not only would this
inexplicable diversity and success of the mammals take an extensive history to
generate these renovations, but they would have to do it under the same
conditions of duress that had supposedly caused the extinction of the
dinosaur.
This makes the relationship between the sudden
dinosaur extinction and sudden wide range mammalian success impossible. Based on logic alone, there are only two
possibilities left for them, and neither is good. One:
this wide range of mammals did already exist, and
therefore the extinction of the dinosaurs was not a catalyst, and mammals had
to survive the same conditions that killed the dinosaurs. In this case, mammals were no more “advanced”
than dinosaurs, having a contemporaneous appearance, and the application of the
evolution scheme is unjustified. Or two:
there was no more than a sparse population of small mammals, like the shrew,
and as dinosaurs were being extinguished, the mammals’ genetic mutations
suddenly exploded into a fever of creativity at such a rate, under these same
catastrophic conditions, that it left no evidence.
Since numerous fossil sites in the world place
dinosaurs contemporaneously with mammals, the causational relationship is
unfounded. Though evolutionists always
interpret such overlap as being the Late Cretaceous, mammalian fossils buried
in the same strata as dinosaurs rationally eliminates the impetus for the
“explosion” in mammalian evolution since clearly they already existed. For example, in
A Guide to Dinosaurs recognizes the
co-existence of mammals and dinosaurs on page 55:
According to the fossil record, these groups,
already diverse in the Cretaceous, later became even more diverse, with large
numbers of mammal and bird lineages appearing early in the Tertiary.
Since fossil evidence shows that the two groups
did live contemporaneously in diversity, we know that they both would have
suffered from the disaster. There is no
evidence of a relationship between the extinction of the dinosaurs and the
evolution of this great mammalian diversity.
The entire evolutionary assertion of progress to higher animals becomes
an illusion if we cannot effectively separate the timeline for the dinosaurs
from the mammals. Considering that the
little shrew would have to radiate out into all these families so suddenly, it
is illogical impose an impossible fast-track evolutionary process that would
produce a hoofed caribou from a shrew about the same time that the shrew felt
pressure to evolve just as abruptly into an anteater.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Interpreting the Fossils
The class to class, dinosaur to mammal evolution
that scientists impose on the fossil record is a narrow interpretation not
actually dictated by the evidence, but formulated through paleontological manipulation. The “geologic column” (as will be discussed in
more detail in the Geological Evidence section) does not line up, one age on top of another, and
so on. One
will not find these perfect layers of strata in representation of this
“progression” of life that contain reptiles in the lower level, then dinosaurs,
then dinosaurs and small mammals, then only the mammals. Generally, only one “age” of rock is exposed
at a time, and a paleontologist will designate the age of the strata based on
evolutionary expectations, and will never allow for an age to demonstrate
contradictory evidence.
For instance, if a dinosaur is found in the
sediments, then the age of the strata will be placed before the Tertiary. If an “advanced” mammal is found, it will be
placed in the Tertiary and later. If a
small mammal is found in the same proximity as a dinosaur, then that age will
be placed at the borderline between the dinosaurs and the mammals, in the late
Cretaceous. If a large mammal is found
on the same level as a dinosaur, than the Paleontologist declares that the area
has been “reworked,” meaning that some geological phenomenon, or erosion took
place, whether detectable or not, to put them on the same level.
One good example of this “reworking” of evidence
is found in the Atlas of life on Earth, page 225 about some large
The unwary creatures may have lost their footing
and fallen down a sinkhole, to be found when the Bernissart coal miner tunneled
through the Carboniferous rock 135 million years later.
Since the miners clearly were digging for coal
through solid rock, evidence of this mysterious, dinosaur herd sized sinkhole
has been imposed on the evidence.
Otherwise, they would have to admit that the evolution scheme is nonsense,
and the dinosaurs were buried in these vast sediments because of the Flood.
Evolutionists are very creative in explaining
misplaced fossils. Thousands of dinosaur
fossils have been found in the exact same sediments as “modern” mammals in
regions around the world. They are never
placed in the same age if doing so would disrupt the evolutionary timeline no
matter how close the proximity, or how similar the strata. Therefore regardless of what the evidence
dictates, a dinosaur will always be placed in its “proper” age, and a mammal
will always be placed in its “proper” age, and never the two shall meet.
Because of this, the age placement process for
fossils is based on evolutionary circular reasoning—“Mammals didn’t fully
evolve until after dinosaurs because we don’t find any advanced mammals in
earlier strata, therefore any mammals we find must be from later strata.” Evolutionists simply will not allow
“advanced” mammals and dinosaurs to co-exist.
Although they are responsible for how fossils
are dated within their own scheme, they actually turn this around and argue
Creationists with “Why then don’t we find “advanced” mammals on the same level
as dinosaurs?” Well, for one, the
“advanced” status is arbitrary; two, we do find them on the same level, though
you won’t acknowledge it; and three, it is uncommon because evolutionists don’t
have to place them on the same level when there are so many ways
to avoid doing so.
In fact, even if an “advanced” mammal, like a
badger, were found in the death grip of a dinosaur, (which is highly unlikely
since they were drowning at the time) paleontologists could simply call into
question whether or not the badger was a true mammal, or instead a mammal-like
reptile. Perhaps the site was reworked,
and some tectonic activity brought them together. If one considers the reality, even as common
as fossil finds are becoming, there is still generally enough space between
them to allow for a multitude of rationalizations. The only reason small mammals are
acknowledged so early in the record is that 1) they are necessary for tying
mammal-like reptiles to mammals, 2) they are found in abundance everywhere, so
they are hard to ignore, and 3) paleontologists simply call them “primitive,”
and feel good about that.
If, however, there were a flood today, how many
different “higher” species of animals would likely be buried side by side? How many foxes would be buried next to
beavers? Though they live
contemporaneously, it would be very rare to actually stumble upon them like
this. The ratio of “higher” mammals per
acre is very low, since they each require a large area for ecological
support. There are only so many lions
per square mile, so having one buried side by side with a T-Rex is
statistically low. Actually being
located on easily exposed strata reduces such occurrences even more. Therefore, though these inexplicable finds do
occur, their low frequency facilitates the ease with which paleontologists can
quietly dismiss them.
Different geologic ages are frequently assigned
to strata that are obviously uniform in deposition just to avoid a
contemporaneous association of species from different ages. Despite these efforts, there is still
undeniable evidence that clearly shows that mammals were buried in the same strata
as dinosaurs.
Recently, rich fossil deposits have been
discovered in the Gobi Desert, Mongolia, which demonstrate that dinosaurs and
mammals overlapped in abundance. These deposits have offered
at least 187 individual mammals since the 1990’s, in a region where
dinosaurs, crocodiles, turtles, and lizards, have also been found. This Flaming Cliffs area is known for the
extensive dinosaur fossil finds, as well as some famous finds which will be
discussed in the geological section on fossils.
The fact that an area rich in dinosaur finds is
associated with such an extensive mammalian find tells us that there is little
to separate them. Especially when one
considers that the sediments they are buried in appear uniform from the top to
the bottom through any imagined ages.
Astonishingly, scientists have dated these diverse finds to the
Cretaceous rock, about 15 million years (according to evolutionists) before the
extinction of the dinosaurs. This large,
unexpected collection of mammals demonstrates how extensive they were already
in relation to the prime of dinosaur dominance.
Such an early mammalian diversity affords no catalyst for their
explosion, and no rationale for how these varied creatures survived when the
dinosaurs were extinguished.
The conflict between the evidence and evolutionary
timelines is not uncommon. Sometimes, it
is unavoidable, but evolutionists refuse to bend to the implications. A Guide to Dinosaurs illustrates this
willing denial on page 217 concerning the finds in
The controversial occurrences of dinosaur
remains within Paleocene deposits at Hell Creek has led some paleontologists to
suggest that dinosaurs could have survived the meteor impact and lived on for a
short while in the early part of the Paleocene.
Others, however, interpret these occurrences as being nothing more than
the re-sitting of Cretaceous remains into Paleocene layers as a consequence of
the erosion of the Cretaceous deposits.
The entire evolutionary scheme falters under the
interpretation that dinosaurs lived after the great extinction. This is the very event, which necessary for
explaining the sudden explosion and diversification of the mammals. Here, evolutionists must resort to altering
the natural conclusions about the evidence, and impose conditions that are not
evident in order to maintain the scheme.
Moreover, even these concessions are limited without a cause. If dinosaurs are found in the same strata as
higher mammals, then there is nothing to support the notion that they were ever
separated by ages, or that evolution operates at all.
The assumptions about both the extinction of the
dinosaurs and the success of the mammals are built on false premises. The fossil evidence does not suggest that
mammals and dinosaurs definitely lived in compartmentalized ages, but it
confirms that they definitely coexisted, and with diversity. More on the topic of incongruence in the
fossil record will be discussed in the section on geology.
Such compartmentalization is not evident, but
imposed on the evidence since both are frequently found in the same regions and
in related strata. Fossil dinosaurs labeled
from the Jurassic-Cretaceous are found in abundance in the Southwest and
Midwestern United States, in the same region and sediments as a profusion of
mammals placed in the very near Tertiary, with only the thin veil of assumption
separating their ages. Therefore, because paleontological timelines are built
upon this assumption that mammals excelled in the wake of the dinosaur
extinction, there is no other evidentiary basis for continuing to accept the
timeline at all. The fossil record does
not actually confirm evolutionary assumptions, but it is a snapshot of life,
frozen in the sludge of the Flood.
Evolutionary scientists from all fields generally concede that there are
no fossils in the vast record to demonstrate how bacteria turned into
invertebrate, then an invertebrate into a fish, and then a fish into a land
dweller, into dinosaurs and mammals over millions of years, despite the fact
that there are sometimes thousands of feet of suitable sedimentary rock
available to preserve them in.
There is no other way of proving
that evolution really happened except through the history that fossils
tell. If there is no verification of
transitions in the one place that evolution ought to be most evident, then
evolution is not the most reasonable interpretation. When a scientist seeks to explain
why he has not found the evidence necessary to support his theory, and why
that evidence appears to support the opposing theory, but uses no factual
evidence to do so, then the unbiased perspective necessary for scientific
research is replaced by a belief system that one is seeking to substantiate.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Shared Traits
The fossil record clearly demonstrates that all
species are in stable forms, and that there is actually a diminished variety of species
today than in the past. The fossil record also
establishes that species living today would have remained remarkably unchanged
over millions of years of “evolution” until the present. The expected idea that millions of years of
evolution has climaxed in all the great diversity of life today, is not
demonstrated in the fossil record. It actually shows us that
life has always been complex, and diverse, but a great deal of this wonderful
variety has been lost, not gained (for instance, we are missing a lot of cool
marsupials).
Nothing in the record indicates that today’s
life is more developed or more complex than the past. In fact, the lower forms thought to be a part
of the evolutionary climb toward higher forms, are generally still present in
today’s living world, testifying that it was not necessary for them to go
through all those changes to survive.
Every inexplicable explosion in animal development immediately settled
into forms that would remain stable until this day. What motivated the little bacteria that had
the world to itself, to become a human when that bacteria today is as stable as
always? Perhaps, even bacteria can
dream.
Since there is no actual fossilized evidence to
document the transition of species, there are numerous questions that
evolutionists are hard pressed to answer.
One issue is the fact of shared specialized traits among different
animal classes, which would have developed on unrelated evolutionary branches. Evolutionists call this Convergent Evolution,
because these traits would have had to form separately in unrelated
species. Convergent Evolution cannot
explain the miracle of traits shared by unrelated classes—it just gives it a
name.
A terrific example of a problematic shared trait
is that both the animal world and the plant world operate on the exact same
system for multi-celled sexual reproduction, by meiosis. This is a complex feat that is hard enough to
explain the first time. Considering
animals and plants would have evolved into multi-celled structures independently,
it is more remarkable to do it twice. Multi-celled plants and
animals could not have a common multi-celled ancestor, and therefore, they would
have coincidentally invented the same process during the course of their
haphazard evolution. Since animals
theoretically pulled it off first, maybe the plants got jealous and copied
them.
There are a great many examples of shared traits
among animals. A major example of this
is found in wings. There is no
evolutionary explanation for winged flight successfully developing among
unrelated animal classes other than through extraordinary providence. Every animal class has an example of
flight—insects, mammals, birds, fish, and reptiles. Volant (flying) reptiles are found in the
fossil record as Pterodactyls and related species. There are many species of flying fish, which
can actually fly for hundreds of feet, though being fish, they do not remain aloft
for long periods of time. Volant mammals
are represented by the abundant species of bats.
Now there is no suggestion made among
evolutionists that all these species have branched off from a common
ancestor. There is no implied link
between insects and Pterodactyls, or birds and bats. These species would have evolved on
completely separate paths, yet they share a remarkable feature that
unbelievably would have appeared and developed separately through millions of
genetic mutations, without any idea where they were going. The mechanics required for a body built for
flight are so incredible that it completely discredits the possibility of them
developing in any animal accidentally even in one species, but to do it in
numerous unrelated animal classes is absurd.
More critically, this marvelous development of winged flight is
completely undocumented in the evolutionist’s only hope for evolutionary
evidence—the fossil record—as each species appears suddenly and fully
formed.
Wings are just one example of shared traits
among animals. There are numerous such
coincidences among the different classes.
Jointed legs exist in
arthropods (bugs), and crustaceans (crabs) through one branch, and birds and
tetrapods. The complex hard-shelled eggs
are produced by both birds and the Duckbill Platypus without a common
ancestor. Warm-bloodedness and related
metabolic features are shared by mammals and birds, though they supposedly
took separate evolutionary routes (birds through dinosaurs, mammals through
reptiles). Since birds and mammals would
have come through different classes, this complex metabolic transformation
would have had to evolve twice.
Cold-blooded reptiles would have had to evolve warm-bloodedness for
mammals, and dinosaurs, which came from reptiles, were also likely cold-blooded
before they would have evolved into birds.
Some evolutionists propose that dinosaurs could
have been warm-blooded, but this is unlikely for many reasons. Warm-blooded classes of animals have a fur, hair,
or feather covering, or at the very least, blubber and the like for heat
regulation. Since no classes of animals
without these insulators are warm blooded, then the scale covered dinosaurs (according to the scaly skin
imprints from inside shells and other fossils) confirms that they were more likely
cold-blooded. Additionally, like
reptiles, scales do not contain pores, which would indicate the inability to release
heat through the skin, in addition to a lack of sweat glands.
Regardless of whether the dinosaur was
cold-blooded, mammals and birds still would have come through different reptile
branches, necessitating the unbelievable new metabolism to be invented
twice. Remarkably, birds and mammals also share
a four-chamber heart, which reptiles and dinosaurs don’t. Additionally, birds and mammals both have follicles
in their skin in order to produce hair and feathers, although they supposedly
took separate routes from the reptiles, which have scales, a completely different cell structure incapable
of producing hair or feathers. Even
insects and arachnids have hair on their bodies.
Further examples are that amazingly both whales and bats have
the complex echolocation system, though they are quite separate on the supposed
evolutionary path. Even the swiflet bird of
Remember also the complex eye of the squid and
the complex eye of the human, which would have no common ancestor, are almost
identical in structure. Squids most certainly did
not evolve into fish, so fish and all their fishy decedents had to have
re-invented the marvelous eye. Again,
powerful evidence against these tall tale notions of evolution is that the
squid (or nautiloid) is supposed to be among the first multi-celled organisms
produced by the Cambrian explosion, and yet it had already fully developed the
remarkable eye by this supposed earliest stage multi-celled life.
These examples are just a few of the remarkable
features shared among unrelated branches of the evolutionary tree. In fact, one of the greatest mysteries to
“evolution” is found in marsupials.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Marsupials
One of the most astounding examples of shared
traits is found between marsupial and placental mammals. Marsupial fossils have been found throughout the
world, and demonstrate an
astonishing variety. Although we are
most familiar with
The unique method of birth in a marsupial sets
it far apart from placental mammals. Physiologically, the two are not
interchangeable, so evolutionists readily agree that this phenomenon developed
separately from placental species. The
problem with this is that the only shared characteristics among marsupials are the physiology,
method of birth, and in most cases a pouch for the young—otherwise, there are
currently about 250 marsupial species, with hundreds of fossilized extinct
species. The majority of these species possess
otherwise nearly indistinguishable body types from their placental
counterparts. So while the marsupials
themselves are very different, they are uncannily reflective of the placentals
they are not related to.
Examples of shared body types with placentals
are marsupial versions of mice,
rats, shrews, anteaters, moles, wolves (recently extinct), cats, and
squirrels. The placental wolverine and
the marsupial Tasmanian Devil are good examples of matching body type. Most remarkably is the resemblance of the
Southern Flying Squirrel to the marsupial Sugar Glider. These two flying “squirrels” are so identical
that the non-biologist would be easily confuse them. Their gliding features mirror each other
precisely, yet they would have evolved independently in these two unrelated
subclasses.
The similarities in body type are too remarkable
to ignore even between placentals and the more unusual marsupials, like Koalas,
wallabies, opossums, and Kangaroos.
Marsupials exhibit all the familiar mammalian features so that most
people cannot tell marsupial mammals from placental mammals without someone
identifying them. Some of these basic shared
physical features include fur, whiskers, claws, type of teeth, facial
structures, basic skeletal structures, ear structure, warm-bloodedness,
metabolic functions, mammary glands, live birth, and innumerable other common
mammalian features. There is no question that
these two types of mammals resemble each other, have the same metabolic
functions, and live in the same “types” of habitats.
Evolutionists are faced with believing one of
two remarkable things about marsupials.
One is that through miraculous coincidence, each of these unrelated
placental mammals (mice, cats, wolves, anteaters, squirrels, shrews, moles. .
.) independently each developed this completely unique marsupial method of
birth. This means that each species
coincidentally reinvented marsupialism scores of times, and produced
counterparts for placental mice, cats, wolves, anteaters, squirrels, shrews,
moles, etc. . . The other option is that
the marsupial branch and the placental branch split from each other way back in
the earliest mammalhood some time in the Jurassic, and marsupials, in the
course of evolution, coincidentally diversified into this wide variety that so
precisely mirrored their placental counter parts.
Marsupial birth and physiology is so unique that
evolutionists generally choose the latter, and assert that placentals and
marsupials emerged from separate evolutionary paths. This is a damaging concession, since it
allows so much diversity in mammals prior to the demise of the dinosaurs—and
yet there is still no fossil evidence to support the assumption. The more pressing question that cannot be
answered by the fossil record is how did the specialized marsupials, splitting
off so early from a common primitive ancestor, manage to perfectly imitate
their unrelated counterparts?
One may wonder how, by luck, evolution produced
a placental anteater, and a
marsupial anteater. It supposedly took
millions of years for mammals to develop, yet somehow placentals and marsupials
took closely parallel contemporaneous paths without any impetus in their
genetic makeup to do so. This is a
completely unreasonable and unscientific notion, and the parallelism of
marsupial and placental body types is a fact that refutes evolution, and
supports Special Creation.
Like marsupials and placentals, there are dozens
of animals with shared traits that are so far from having an evolutionary
explanation, but evolutionists refuse to acknowledge the implication of these
impossible examples. Not only does
Creation mock scientists with such unusual montages as the Duckbill Platypus,
but even common animals defy evolutionary explanation. Although these species clearly share traits
with other species, no fathomable lineage can be erected between them. They just hang out there dangling off of
little dotted lines from the evolutionary tree.
For example, despite their worldwide success,
there is not a single fossil that even implies any link between all other
mammals and bats. They are utterly isolated on the evolutionary tree, and fully
developed from the earliest fossil record, which has preserved an abundance of
specimens. Scientists can’t even begin
to demonstrate how they came to be, and yet there are thousands of bats in the fossil
record, and nothing but complete bats. Even
the imagination is unable to fill in the millions of years of physical
transitions necessary to produce these remarkable creatures. Despite the fact that insect, rodent, and fish
eating bats usually have poor vision at night, they can catch fast moving prey
through their complex echolocation system more effectively than most animals do
by sight. There are no fossils, however, to link them
to the rest of their “family tree.”
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Whales and Dolphins
Another interesting example of shared traits is
found in marine mammals. Evolutionists
believe that pinnipeds (seals, sea lions), sirenians (manatees) and cetaceans
(whales and dolphins) had to arise through different, unrelated evolutionary
paths. Despite this, their forms are
extremely similar. Manatees and whales have
broad tail flukes for propulsion, and all three groups have streamline bodies,
flippers, blubber, and revolutionary oxygen preserving physiology. It is remarkable that evolutionists believe
that three different ancestral mammals had the mind to evolve into such
specialized, parallel forms in order to live in such a foreign
environment.
There is no agreed upon link
between supposed ancestral land mammals and these marine mammals. These marine mammals
appear suddenly in the fossil record, at the Tertiary level, fully formed,
without a traceable indication as to their evolutionary pathways. The concept of marine mammal evolution poses
great problems for evolutionists, since mammals would have had to first develop
on land before returning to the sea.
Despite this requirement, marine mammal fossils
are found in the same age, the Eocene, as their supposed ancestors. This causes the evolutionary timeline to
become extremely condensed and indiscernible.
Because there is no clear evidence indicating how such a body revolution
could have happened so precisely and so rapidly, evolutionists must first
attempt to formulate the most likely scenarios that could have brought mammals
back to the sea, and then look for suitable candidates that will best piece
together this series of unlikely transitions.
As remarkable as all marine mammals are, whales
are particularly specialized.
Accomplishing such an unscripted evolution would be an implausible
task. Try to envision the self-sacrifice
over millions of years of these valiant, hairy, four-legged creatures,
determinedly immersing themselves into the water until their genetic mutations
caught up. To witness such a transition
would be remarkable, and surely one would think evolution was going the wrong
way. Many transformations would be required in
order to make a scrawny, four legged land mammal into a robust, yet gracious,
streamline whale. It would be necessary
to remove its legs, abundant hair, move the nostrils to the top of the head,
provide complex echolocation features, streamline the body, invent fins and
tail flukes, rubberize the skin, and produce blubber for insulation and
buoyancy, create floating stones in the ears, restructure the eye lenses for
seeing under water, greatly modify the spine and cranial structures, multiply
the size of the creature numerous times, and add innumerable other water
dwelling features.
Additionally, whales give birth backwards, have
squirting milk mechanisms, and newborns are able to seal off their esophagus
when nursing, all so they won’t drown (these features would have to evolve fairly
rapidly). In order to perform deep sea
dives, whales are also equipped with highly oxygen absorbent muscle tissue, and
much more oxygen absorbing hemoglobin is present in their bloodstream than in
other mammals, along with collapsible lungs and other depth adjusting features. More fascinating is the peculiar path that the
blood takes to the brain of whales through blood vessels associated with the
spinal cord instead of through the carotid arteries like in other mammals. These particulars don’t
include all the other incredible physiological and skeletal adjustments
necessary in evolving into a whale, as the only real resemblance between these
creatures and other mammals is scarcely noticeable to the average observer in
comparison to the incredible differences.
These poor creatures supposedly endured all this
painfully awkward evolution because it was the better path to
survival, while the other lucky mammals stayed on the dry land, effortlessly
continuing to flourish in the environment they were suited for. What motivation they must have had! What
determination! And they didn’t even know
how it would turn out. It is a mystery
how the intuition of the mammal and the fortuitous genetic mutations could have
worked together toward such perfection.
One would imagine that such a drastic transition, taking millions of
years, would be able to yield numerous clear fossil pathway links, but though
there are numerous whale fossils, and numerous terrestrial mammal fossils,
there is nothing that can practically represent any intermediate species.
Whale evolution, let it be said, is
impossible. Do not let the existence of
this discussion serve as some form of acknowledgement of its validity. Evolutionists have so nothing to work with in
the fossil record, that they substitute assertions for facts until no one
questions it. “Whales evolved, that’s
it—we just don’t know how,” they say.
Some scientists speculate that a bovine-like creature may have been the
lone adventurer.
Many others are mysteriously convinced that it
was a carnivorous wolf-like creature called Mesonyx, which resembles a whale in
that it has an
axial skeleton
(like all other vertebrates). It also has hoofs. One would never guess that this dog-sized,
land dwelling carnivore was the first step to whalehood, but apparently the
famous evolutionist Stephen J. Gould was certainly convinced. He stated in the Natural History Magazine,
May 1994 edition, that he concurred with the transitional series assumed to be
true at the time, which went from Mesonyx to whale through the new find,
Ambulocetus—meaning walking whale. He
thumbed his nose at Creationists in triumph, saying:
If you had given me a blank piece of paper and a
blank check, I could not have drawn you a theoretical intermediate any better
or more convincing than Ambulocetus.
These dogmatists who by verbal trickery can make white black, and black
white, will never be convinced of anything, but Ambulocetus is the very animal
that they proclaimed impossible in theory.
Now, unfortunately for the enthusiastic Mr.
Gould, the official status of Mesonyx has been tossed into confusion by Philip
Gingrich’s research. Gingrich, who had
himself always maintained that Mesonyx was the ancestor of whales, became
convinced otherwise by his own finds in 2001.
Gingrich believes that his discovery of Pakicetus is certainly in the
lineage of whales, and he has had to adjust his thinking about Mesonyx. Why?
Because according to his analysis, these are the first whales ever
discovered with the proper ankle bones found intact. That makes sense, right?
But to everyone’s surprise, these early whales
had ankle bones like the family that cows, sheep and hippos supposedly came from
(artiodactyls). So now, whales are
joined to this unique family, and Mesonyx must be quietly discarded, making
poor Mr. Gould appear a little premature in his assessment of the perfect
transition. Never fear, though Mr.
Gould, because although Ambulocetus doesn’t have the right ankle bones to
reflect a transition from a sheep-boned ancestor, apparently evolutionists
don’t mind, for they seemed to have decided to keep this four-legged creature
in the lineage. See how easy that
is? You are a scientist, so you get to
say things like “This was the perfect transition between these two animals, and
now it is between these two very different animals too.” And you can say “This four legged animal is
the first whale with the right ankle bones,” and thus it is so.
What is so stimulating about these transitional
choices is how baffled the researchers themselves are by the evolutionary
puzzle. In the
They’re so different from other warm blooded,
furry things that it’s been a mystery, both how they came to live in the sea
and what ancestors they might have come from on land.
Yes,
quite a mystery. Interestingly,
researchers are just as convinced now that whales are related to sheep and
hippos as they were that they were related to hoofed wolves prior to that. Especially since the DNA comparisons have
always pointed to a more hippo/sheep relationship than the wolves. Meaning, when they count all the little
places that DNA shows up in the same general sequence, more of these places are
common between whales and this hippo/sheep family than between whales and any
other family of mammals.
Now when one assumes that whales must have come
from some terrestrial family, one must settle for what one can
get. So now we have Pakicetus, a
wolf-like, sheep-family land whale. Of
course one problem with this new assumption is in imagining the motivation for
any sheep/hippo/cow/deer related creature to enter the sea. That would seem the wrong place to go for
such a creature since they are all herbivores and there is nothing in the ocean that they
would like to eat. Whales, as we all know, are
carnivorous, and eat animals. Pakicetus doesn’t look much like a
sheep or hippo at all, and perhaps it was a carnivore, but then one
wonders if it really is related to the sheep family, as desired, after all.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
One
constant bit of frustration about whale evolution is the amount of
inconsistencies associated with it.
There are many minor cetacean candidates vying to fill the charts, but
they are disturbingly difficult to track. One commonly reproduced diagram from Evolution,
the Triumph of an Idea by Carl Zimmer shows one mysterious candidate,
Rodhocetus, as whale-like and without notable external legs, but another text
says that it has a full set of powerful hind legs and likely could go on
land. Another shows Pakicetus walking
normally, and the next claims it was seal like.
One depicts Ambulocetus, and the next shows that same depiction labeled
as Rodhocetus.
These inconsistencies can be found throughout
the journals and textbooks, on major levels.
It would be wise not to accept any characterizations of fact in any form
until it is either found common among the majority of sources, or the actual
bones themselves are pictured. One may
never know if the picture is labeled correctly, but it would at least be
possible to detect whether or not there were legs.
Perhaps the confusion is caused by the
evolutionists’ crafty characterization of the fossil finds. Not desiring to leave us to draw “the wrong” conclusion,
textbooks and papers frequently blend the facts in with the imagination so one
cannot tell the two apart. The
confidence inspired by such specimens as Rodhocetus, Pakacetus, Ambulocetus
(and whatever else might come along) does not betray the fact that none of these
candidates have been found with their pelvis, back limbs, and tails even
remotely complete. They are, however,
all four-legged.
Scientists would like us to believe that they
can magically divine what the entire lower half of an animal looks like based
on the upper half, even down to whether the feet were webbed or not. That would be nice. They take it upon themselves to help our
imagination, and draw entire depictions for us, even describing features that
have not been found, but inferred.
Moreover, they do not have the forethought to agree upon the major
details that they construct, and often contradict each other, although all are
equally certain. Whenever the rest of
the animal is eventually found, and the original conclusions don’t hold up,
rather than publicizing the error, and learning from it, the traditional
response is to confidently move on, in the same manner, to the next possible
assumption.
In fact, since Gingrich’s first conclusions, more of
Pakicetus’s post-cranial remains have been unearthed, including the pelvis and
hind limbs. While initial assessments of
the partial Pakicetus depicted it as a swimmer, all studies of these new finds
have caused researchers now to conclude unquestionably that the animal was built
for walking on land in the normal manner.
But they still, bafflingly, refer to it as a whale.
The matter of exactly how the whale evolved from
land mammals is still arguably undecided, (but nonetheless absolutely true).
Whale evolution has become a self fulfilling paleontological prophesy that
begins with a conclusion, and ends with an assumption. Philip Gingrich himself characterizes his
other finds in
Our 1977 fieldwork in marine strata yielded more
archaeocetes (ancient whales),
which foolishly we were not interested in at the time. Pelvic bones that we found that year were
attributed, questionably, to land mammals because it was impossible to imagine
that whales had such robust hind limbs.
Apparently almost 25 years later, after little
success in finding feasible transitional whale species, his imagination became
more pliant. Gingrich’s assessment of
finds has lead him to conclude that these proto-whales used their powerful back
limbs, sheep-like ankles, and elongated feet as the main means of propulsion in
the water. Perhaps he believed that this
powerful kicking behavior persisted until its legs very dramatically dropped
off in favor of the flukes and tail mode of propulsion.
Such a process is not very compelling evidence
of any relationship to whales, though, since the investment in powerful hind
legs is in no way a move toward whaleness, but perhaps toward giant otterness. Even otters don’t rely so
much on the power of their legs for rapid swimming, but rather they undulate,
and assist with their feet. However, it does not appear that otters are
interested in giving up their legs. Gingrich’s assessment that those powerful
legs would have been the best source of propulsion would be just like
the—actually, like nothing. No animal built for swimming swims with its legs as the
primary propulsion. What, the duck? The polar bear? That’s it.
Gingrich must be picturing the future whale swimming like a polar
bear. Hmmm . . .
There are many animals that are equipped for their
environment, and are capable of doing marvelous things, but they are clearly
not related to whales. There are
scavenger hyenas (Brown hyena) that eat along the shores, and there is a hoofed
animal that can get by at swimming a bit (the deer-like Sitatunga of Africa),
and a few furry, four legged mammals that can swim and catch fish (the otter,
polar bear, and fishing cat). There is
even the great hippopotamus, which is a giant, graceful four-legged loper more
whale-like than all the rest, but it walks (not swims) on the bottom of rivers,
and is no more related to whales than a sheep.
Although all these species can function in
marine environments, these living animals aren’t even in the same genus with
each other, let alone whales. The scant
presumed whale candidates have no more of a legitimate link to whales than
these animals do. When we take into
account the capricious nature of transitional offerings, we could be looking
for a wolf or a sheep—perhaps a wolf in sheep’s clothing. Once there are enough options out there,
evolutionists will not actually have to tie them together in any rational
progressive order.
This is what we have so far. Mesonyx has been discarded for having the
wrong ankles. Pakicetus has the right
ankles for a whale, but it is fully terrestrial, so it is an entirely
subjective first candidate. Then
Ambulocetus, Rodhocetus and all the other little cetuses have all four, very useful
legs. They are no more than 11 feet long, and do not inspire a
sense of whaleness at all (but their heads are kind of big). Now what we can do is ignore all of those
problems and just assume that if a whale evolved from a terrestrial mammal, it
had to start somewhere. Let’s simply
allow evolutionists to fuss around with these four legged animals since they
are still trying to figure it out, and examine the next nearest transitional
candidate.
Now the next nearest link to these small,
four-legged creatures is Basilosaurus, which is a serpentine monster, with miniature hind
legs, that grows to a whopping 65-80 feet long. There
are no intermediary candidates closer to the previous candidates than
this. This creature, quite unmistakably,
is a great jump in both size and form.
It is a lot like evolving from a mouse to an elephant in one genetic leap.
Textbook illustrations frequently depict these
various candidates as if they are the same relative size, despite the difference of up to ten
times the size between them and Basilosaurus. It
would be hard for the logical mind to accept such a disparity with out gradual,
intermediary transitions. There is
another fossil candidate called Dorudon, but Paleontologists put them in the
same family as Basilosaurus, and other than being only about 20 feet long, there doesn’t appear to
be a marked difference between the two.
One might conclude that Dorudon is just a Basilosaurus not fully grown
since admittedly, no baby Basilosauridae have been found.
Despite evolutionists’ grandiose enthusiasm for
whale transitions, it is plain that there is actually a great void between the
small four-legged “aquatic” animals, and the next best thing to a whale, the
gigantic Basilosaurus. Although the
early candidates from land to sea are furnished with desperate hope,
Basilosaurus is the first candidate to at least exhibit some appropriate
aquatic features similar to whales.
However, at this point, only a few paleontologists still accept
Basilosaurus as a whale ancestor. And
actually, a growing number of scientific texts are removing Basilosaurus out of
the whale lineage and relegating them to a side branch because of the
difficulties we are about to discuss.
This concession alone by the majority of paleontologists was the last
proposed link between land mammals and whales, leaving nothing to establish its
evolution but vain necessity.
Basilosaurus is one of the few actual fossil
species included in whale evolution that is known to be complete and
abundant. As mentioned, it is the
closest paleontologists can get to whales, and it is not that close. It is an enormous, serpentine creature that
clearly lived in the water. Besides the
lack of transition from previous candidates, there are many problematic
features that would complicate Basilosaurus’ evolutionary ancestry to whales. There are no cranial developments that
whales have in order to perform their special diving functions. Overall the skull and jaw structure is not
analogous to whales, and the teeth (except for the front few, which are pegged)
are triangular and serrated for powerful chewing of flesh.
Toothed whales and dolphins actually have all
pegged teeth with a similar function as alligators that only grip, and do not
slice or chew (the way that terrestrial carnivores do) because they swallow
their food whole. Additionally, the
shape of the skull does not allow any development in the echolocation “melon,”
which dolphins and most toothed whales have. Evolutionists simply have no pathway from
terrestrial mammals to baleen whales (which is another in depth, but no less
pointless issue), so Basilosaurus can only connect with toothed whales.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Any legitimate proto-whale must possess features
that suggest adjustments for deep diving and echolocation. Basilosaurus does not possess any skeletal
advancement in this feature. Most dolphins and many toothed
whales have a “melon” organ that gives whales this ability. The toothed whales that do not have a melon
organ still produce echolocation with sacks and muscles by their blowhole. Basilosaurus, however, did not have a
blowhole, and therefore breathed through its nostrils in the normal manner.
Additionally, Basilosaurus doesn’t have the
palate, sinus, or ear configurations seen in whales necessary for basic diving,
breathing, and echolocation functions. Cetaceans require these
structures in order to perform the deep diving, and lengthy submersions, and no
progress is evident in Basilosaurus. Even paleontologists admit that
Basilosaurus probably had to stay near the surface. Cranially, the two share only
superficial similarities, but Basilosaurus could not begin to perform these
basic whale diving functions, and there is no available next candidate to
bridge the gap.
Basilosaurus also has small, but unusual hind
limbs. Creationists agree that they
would not be of much use on land because they are so infinitesimal. One would think this is a good argument that
the legs were on their way out. The
problem is that even as small as they are, they are well developed. They are clearly functional with all the
necessary parts: pelvis, femur, patella, tibia, fibula, ankle, foot, and toe
bones, complete with groves for the ligaments. Yet
they are so small.
Evolutionists take these small limbs and apply a
number of adjustments to help their theory.
Some completely detach these little limbs from close proximity with the
spine in order to facilitate the illusion that Basilosaurus has a very broad
body like a whale. When the legs are
depicted in near articulation with the spine (the way it was found), then the
animal is clearly serpentine because of the narrowness of the body.
Some evolutionists internalize the legs and call
them vestigial—like a heap of useless bones encapsulated within the
animal. This is great if you want to
claim that they correlate with the very useful hip bones found in whales
today. But the elongated scoop-like hip
bones in whales, which evolutionists would like to say are vestigial, are
simple anchors for the reproductive organs, and do not reflect the remnant of a
complex set of functional limbs. Most
evolutionists agree, however, that these limbs were too complex to be encased
within the body, and their relationship with the spine indicates that the
creature was quite serpentine. The
likely use for the limbs was to aid in copulation.
Moreover, the structure of the elongated spine
and tail are clear evidence to paleontologists that this creature did not have
the robust, specialized vertebrae necessary to support the powerful
flukes. Whales have special vertebrae
for these massive muscles and their jet propelled up and down tail strokes. The body of Basilosaurus is so long, that the
thin spine could not perform this movement for propulsion. Although there are small ball vertebrae
indicating a broadening near the end of the tail, many paleontologists
recognize that structure of the spine is more consistent with a slithering side
to side movement, like an eel. The true
image of Basilosaurus might well be gleaned from the first impression of its
discoverer, who promptly associated it with a serpentine reptile—hence the
“saurus” part of the name.
Whether or not it is a reptile, its serpentine
nature is evident when one is willing to see.
The features necessary to show that it is an ancestor to whales are
undoubtedly missing. Even at what would be considered a late stage in
whale development (in fact, what is supposed to be the greatest, nearest link to
whales), Basilosaurus does not yet begin to approach the specific features that
define a whale. Basilosaurus is missing the
development of the melon and cranial features for diving and echolocation, the
presence of a blowhole or necessary palatal structures, the powerful whale jaw,
or the robust spine to support flukes.
One surprising bit of information is that this
grand evolution from land mammal to Basilosaurus, to whale, supposedly all occurred in
the Eocene—just one geological epoch.
Evolutionists believe it took hundreds of millions of years for the
vertebrate fish to become mammals, but mammals would have rapidly transformed
into these highly specialized marine mammals (undergoing major alterations) in
less than 15 million years. Moreover, Basilosaurus,
despite not having any deep-diving features, has to make it all the way across
the vast ocean from
Despite the insistence of evolutionists that
whale evolution is a fact, the branch is actually completely bare. The excitement generated by four-legged
creatures is ludicrous in light of all that is would take to become a whale. Any scientist that categorizes a creature as
a proto-whale because it has the right ankles is certainly desperate for
anything. No argument about the measure
of an animal’s aquatic life can justify it as a proto-whale if it has four
legs. When the next closest candidate to
these small, four-legged animals is a gargantuan serpentine creature, the
theory is unrealized. Since Basilosaurus
itself has been discarded by most evolutionists as being a slithering, giant,
surface-dwelling non- cetacean creature, then nothing remains of the evidence
for whale evolution.
While we all would agree that there are fossils
of many unusual creatures out there that we do not have today, the problem
remains that whales and dolphins still have no feasible fossil history of their
admittedly rapid and severe evolution.
There is nothing even remotely resembling a series of stages from land
animal to the graceful, specialized whale.
Whales suddenly appear in
the fossil record, worldwide, fully formed, with all their unique features, and
completely recognizable.
As certain as evolutionists are about the
evolution of whales, not one can offer any plausible chain of transitional
species to illustrate the vast transition from land to whale that even most
evolutionists agree with. The reality
is, scrounging for intermediary fossil candidates is even easier than logically
dealing with the implausible concept of a furry, four legged, hoofed creature
insisting on living in the water until he and his genetics got the hang of
it.
Just as in every other proposed evolutionary story,
there is nothing but the sparsest of hope for such a great transition, yet
evolutionists are undaunted. They must
assume that it happened, regardless what the fossil record, and common sense,
dictate because there is no other option for them. Just using the fancy names confuses the
public into accepting the assertion as a fact.
It is not enough to want there to be transitions. In science, ideas should be based on
fact. If evolution can’t explain whales,
then it is an inadequate theory about life.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Animal Conclusion
It boggles the mind trying to visualize a land
mammal forcing itself into the sea to become one of the most remarkable, and
specialized animals in the world. There
ought to be a long trail of clues pointing to these millions of tiny steps,
clarifying to us what is mentally impossible to calculate. This is more ridiculous when we step back and
consider the millions of transitions necessary to create the incredible display
of all life only by means of these cosmic mutations and the
intuitive will of a species as the creative force. These steps would need to rapidly occur
miraculously in thousands of animals concurrently, without any designs to guide
them, in order to drive each species by genetic error further apart, and yet
remain equally successful.
When we examine fossils, our assessments are
generally limited to the obvious visible traits, but biologists know that there
are thousands of less obvious physiological differences between each species,
and these differences could not be represented in the fossil record. Reckless genetic alterations in these precise
systems would keep them from functioning unless fully formed and fully
integrated. The entire body is dependent
on the interconnection of physiological systems, and this notion of gradual change
would, at the least, put incredible strain on the organism. And yet, despite the assumption that these
painstakingly perilous evolutionary ventures are somehow advantageous, the
supposed primitive forms still survive.
We still have the little bacteria, the “primitive” fish, the “primitive”
amphibians, the “primitive” mammal. We
have the same kinds of lizards that did not become snakes. We have the full spectrum of ungulates that
did not become horses, and we have the same kinds of primates that did not
become humans.
We find that although evolution is purported to
be the selection of mutated traits based on the success of that trait in
the species, the great majority of evolutionary alterations would have been
unnecessary adjustments to successful traits.
This process would have caused animals to endure thousands to millions
of years of tenuous physical experiments.
So many thousands of superfluous alterations would have to occur, that
it would be a tedious job to list and describe even those of one
transition. In addition to the unnecessary
transitions, hundreds of dangerous alterations to essential systems and
structures would also be necessary to accomplish the supposed paths of
evolution. How could a species survive
such tampering, when that species would have already been successful? We would know some of these things if there
were a trace of them in the fossil record.
Where are the three-legged, two-headed, one-eyed
creatures in the fossil record? Instead
they are all as beautiful and functional as they are today. What perfect system of oversight weeds out
these flawed experimental designs before they left a record? Did nature intuitively know to select out
some of the whackier designs? How were
the generations maintained through those painful stretches while the genetics
searched for the best body types.
Or perhaps nature always produced the perfectly
functional design every time. That would
be providential. Scientists have
actually tried such experiments on the computer. They designed graphics programs to take basic
steps in evolving a creature to watch its progress. The programs would randomly select from a set
of options for shapes, and movements, and each generation would have a new
random alteration with a specific goal in mind, such as to swim or walk. Scientists were quite proud of the
demonstration of evolution.
However, unlike evolution, the creature was
programmed to evolve, and the options were dictated. This of course is more like an experiment in
design (however chaotic). More telling
is that these virtual creatures never perfected the complex and balanced
designs that fill our fossil record.
They did prove that even pre-programmed chaos would produce long phases
of hideous and impractical creatures, none of which are found in the fossil
record. Even experiments specifically
designed to demonstrate the feasibility of evolution reveal how deficient the
evidence is to confirm it. Despite the
hope that it happened, over and over, the evidence is left to our imaginations.
In imagining these
intermediate species, one wonders at the benefit of risking so much to renovate
them. Some transitions would have jeopardized the full function of
reproductive methods as in the transition from asexual simple cell to sexual
multi-celled organisms, reptilian egg laying to placental and marsupial live
birth, and back to egg-laying (birds and monotremes). There would also need to be the loss of
functional limbs, as in salamander amphibian to caecilian amphibian, lizards to
snakes, dinosaurs to birds (loss of useful front limbs), land mammal to marine
mammal (cetaceans, pinnipeds, sirenians ), and mammal to bat. There would be complete alterations of skin
in all major transitions from invertebrate to fish, to amphibian, to reptile,
to bird and to mammal, and back to marine mammal. There would have been experiments with
respiration (fish to amphibian, land mammal to marine mammal) and tampering
with the spine (fish to amphibian to turtle—there is no fossil history for the
anomaly of the turtle).
Some species alterations needed to experience
reconstruction of the mouth and jaw as in the transition of reptile to mammal,
and reptile mouth with lips, cheeks and teeth to the bird’s solid beak. The change from reptiles to mammals also
would have brought total redesign of hearing systems through major
reconstruction of the ear bones. Even
entire metabolic systems were reinvented like from cold-blooded reptile to
warm-blooded mammal and bird, and heart from two chambered fish, to three chambered
amphibian/reptile, to four chambered bird and mammal. Interestingly, there would have also been a
loss of useful features as in sharp teeth, abundant hair, and a package of tree
dwelling features as in the transition from primate to human.
The incredible fortuity required for even one of
these major changes to be accomplished before each species died of dysfunction
pales compared to the millions of minor adjustments throughout the entire
animal kingdom. Such genetic adjustments
ought to be commonplace even in living forms, but they are not. The fossil record does not validate
macro-evolution, and our experience with living forms refutes it. We have never witnessed the mechanism of
evolution—not even once.
Some species, somewhere, ought to be in
transition, but the reality is that species do not invent
new genetics that adapt to changes in the environment. They die. They
don’t grow long necks to reach the trees, they don’t wander back into the sea,
they don’t develop echolocation, they don’t alter their reproductive methods,
and they don’t grow wings. They don’t
because they cannot tell what they will need in a million years in order to
start on it now, and even if they could see the future, they couldn’t make
themselves a different species. They
would probably just make better decisions.
What we have now, we have always had—lots of
variety, with no ability to integrate or interchange species. We still have highly distinct species, often
with shared traits, and typically with shared designs, but they do not
demonstrate progress. We have bacteria,
and they are not related to squid, we have the hyrax, and they are not related
to horses, we have lemurs and they are not related to chimpanzees. No one is watching and waiting for the sheep
to venture back into the sea, or for a lizard to fly.
What is living now is what was living then, and
we are not confused about the differentiation between species. Why would we interpret the past differently
than the present, when there is no evidence to do so? Shared traits are common in nature—so common
that hundreds of traits are similar among all species, even though
evolutionary schemes would need to place them on unrelated branches. This is inexplicable within evolutionary
scenarios. One can only invoke miracles
so often when claiming to offer scientific explanations.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Plants
In order to complete the examination of the
fossil record, it is important to pick up the discussion of plants once
again. As mentioned in the biology
section, the fossil record reveals that living forms of plants are easily
recognizable even from their earliest appearance. Pressed into the lowest sedimentary rocks, we
find these familiar species, fully formed and fully elaborated, without a trace
of any awkward predecessors. From this
vital evidence, scientists should only draw one logical conclusion—that plants
have always been fully functional and have remained in stable forms as far back
as the record provides. However, despite
this unmistakable fact, evolutionists insist that plants did evolve from
very primitive, and likely unrecognizable beginnings.
Oddly enough, an evolutionist will vehemently
plead that the fossil record is undeniable proof of their theory, even though
the supporting evidence is not actually there.
They will admit to such, and even try and explain the awkwardness of the
situation, but nevertheless, hold tight to its treasures as proof. Even though evolutionists rely on the fossils
as the sole basis for their theory, such evidence neither reveals, nor allows
any transitional period for plants.
Therefore, discussions about the evolution of plants are limited to
speculation because there is no scientific evidence that they evolved. A look at the fossil record of plants is a
confirmation of Special Creation, and stasis.
Biological texts often try to fill in
between plant species as if there are actual fossil transitions in the record
that help map the progress of plants from the water, but there are not. From their first appearance in the fossil record,
an enormous variety of plant life is readily recognized and fully formed—even a
greater variety than today. Each species of plant
seemingly exploded, covering the entire earth at once at their first
appearance, without warning or predecessors.
Mosses, horsetails, cycads,
sequoias, ferns, palms, oaks, willows, sassafras, magnolias and numerous flowering
plants, both living and extinct, are abundantly present, to the point that each
living species is established as complete from its lowest appearance.
One is hard pressed to imagine not only how
large, complex, fully functional plants developed with no evolutionary history,
but also how they came to exist suddenly all over the world.
Knowing that the insect and animal life, often attributed with the
fertilization and spread of plants, was itself in the early stages of
evolution, they could not feasibly be charged with the spreading of plants so
effectively. In fact both flowering plants and
insects have an inexplicably sudden worldwide appearance.
Scientists readily admit that there is no
evolutionary family tree for trees—or any plant for that matter. One textbook, Biology, Exploring Life,
put it this way on page 833.
Because
ancient plants left so few fossils, it is difficult to document the history of
plant evolution.
There are apparently plenty of fully developed
plants in the fossil record, there are just no “primitive” plants to
demonstrate their evolution. Note the
assumption here that, in spite of what the evidence presents, plant evolution
itself is not questioned. The average
textbook chart tracing the progress of plants will have green algae on one end,
and then spokes of dotted lines directly linked to each individual phylum of
flowering plants, conifers, cycads, ginkos, ferns, horsetales, lycophytes,
rhynophytes, and bryophytes, with no transitional species listed in between. Green algae are just not that
impressive. Scientists must concede to
this utter desolation of transitions, which proves that there is no evidence
that actually demonstrates that plants evolved at all.
It should be easy to trace the gradual
development through the fossil record from those “simple” cells, to those
tricky multi-celled plants, with the hard woody structures that stand out of
the water, vascular cells, and all the unique forms of reproduction through the
millions of years of transition. What
specifically motivated them to leave the water?
Could green algae really make enough soil (decayed rock and organic
material) for the whole spectrum of
plants to suddenly invade the entire earth?
What modifications did they make enabling them to become complex,
multi-celled vascular plants? How did
they pull off the feat of sexual reproduction, and manage to replicate the same method of meiosis
that the animals were utilizing, even though they would not share this
evolutionary pathway?
One may attempt to imaginatively generalize
the possible evolutionary paths, but they are not actually noted in the fossil
record. We don’t know how or what
mechanisms caused one plant to divide itself, one plant to produce spores, one
plant to produce bulbs, one to produce seeds, and one to produces seeds through
flowers with fruit, one to be a gigantic tree, and one to be a specialized
parasite. The biological reproductive mechanisms are
completely different, and not interchangeable.
A plant that produces spores cannot cross with a
flowering plant. Yet plants that would seem to be on separate evolutionary
paths can often share common characteristics, such as flowering duckweed, a
flowering perennial, a flowering cactus and a flowering parasite (like the
European Dodder). We would know precisely
how these plants relate if there were any hint of their divergent paths from a
common ancestor in the fossil record, but because there is not, scientists are
willing to use conjecture in the place of facts.
Of all the intricacies of reproduction, flowering plants are the most fascinating, which make up more than half
of all plants at about 250,000 species.
Non-flowering seed-bearing plants (gymnosperms, such as conifers) are
placed in the Devonian system, but according to evolutionists, the remarkable
flowering plants appear suddenly in the strata attributed to the late Jurassic,
early Cretaceous systems, considered to be about 200 million years later. Although sexual reproduction by seeds would
have already been successful for 200 million years, without any warning,
evolution supposedly took this incredible risk in order to produce flowers, a
previously unnecessary feature.
If the fossil record actually left evidence of
millions of years of development, slowly progressing, and spreading gradually
worldwide, then we could have assurance that this evolutionary tale had some
validity. Instead, the sudden appearance
of complex flowering plants throughout the world, without a traceable history, is
a mystery so absolute, one will find little conjecture on it in paleobotanical
discussions. It simply cannot be traced
or explained.
In his letter to botanist Joseph Hooker, Darwin
himself called the sudden appearance of flowering plants in the fossil record
“an abominable mystery,” which still holds true. Consider the two unique structures of flowering
trees and of flowering plants. Supposedly the previous
period, the Triassic, produced large conifer trees, such as sequoias. It is difficult to imagine, then, how the
Cretaceous period magnolia tree (or any of the flowering trees assigned to that
period) managed to develop. Since the
unique woody trunk structure of a tree would have already been established, and
the “evolution” of the flower would have been complex, which one was the magnolia
tree’s ancestor? Did a flowering plant
rediscover the structure of the tree, or did the tree stumble upon the complex
flowering reproductive system—coincidentally at the same time that the
flowering plants were first appearing?
Neither choice is plausible.
Another example is the palm tree and the
cycad. They appear to be perfect
relatives, but the “Permian” cycad is a gymnosperm, and the post “Cretaceous”
palm tree is a flowering plant. In fact,
the palm tree isn’t a tree at all by definition because it is not a woody plant
with a core and bark, but a fibrous structure, so where did it get its flowers from? Or did it start over again too? Now add to these examples a bizarre
compilation like the Galapagos prickly pear cactus that both flowers and has a
large tree trunk, and you’ve got yourself just a few of dozens of such evolutionary
mysteries.
We ought to be able to learn these amazing and
distinctly unique developmental processes for each of the hundreds of thousands
of plants by simply examining the millions of fossil remnants in every era in
the rock strata. But instead the fossils reflect stasis. We can’t begin to tell the story about the
evolution of plants because even the outline is absent from the fossil
record.
Plant structures are unique among living things,
and possess highly complex systems that differ greatly from animals. Envision their woody stems, or their water filled vascular
green stems, their roots, bark, green leaves, internal structures, flowers, how
they breathe, how they eat, how they reproduce, how they grow. Since they are so different from animals, their
developmental histories would have been unique, and yet there is no fossil
conformation of this great biological quest.
Even evolutionists are not satisfied by the
sparse offering of supposedly “proto” plants, which look strikingly like stems
stripped of their leaves. Since the
torrent of a deluge would certainly separate leaves from stems, this is poor
evidence for a “primitive” form.
There simply is no reason why the fossil record
should preserve only the completed, stable forms so abundantly, and omit any
resemblance to an evolutionary path of development. And if they evolved so dramatically in such a
short time, why did they stop after that, and exhibit total stasis for
“hundreds of millions of years” until the present? However evolutionists choose to envision the
progress of plants and their reproductive systems, their theories would be more
convincing if the evidence confirmed it.
Just like the mystery of the eye, one may suppose how part of an eye may
be useful, and one may suppose how plants stumbled upon their remarkable
variety of forms and functions, but the history in the fossils doesn’t confirm,
or allow time for this tedious process to have taken place.
However, the truth about the history of plants has
been recorded in the form of inexplicable “anomalies” in the fossil
record. Because of their complex
structures and characteristic appearance in the geologic column, evolutionists
believe that land plants could not have evolved until about 100 million years after
the initial sea life of the Cambrian explosion. In reality, it is a well documented fact that 60
genera of woody plants, spores, pollen and even actual wood, have been found
out of place as low as the Cambrian level at numerous sites around the
world—paradoxically demonstrating that fully formed forests already existed as
evolution should have been just kicking off.
The presence of pollen at these lower levels alone
shatters the evolutionary schemes, since pollen is a product of conifers and
flowering plants, which should not appear until at least 250-300 million years
later. Cretaceous pollen has been found
in The Holy Cross Mts, Poland at the Cambrian level, and Jurassic plant spores
have been found at the Precambrian level in Ukraine—ridiculously impossible
for evolution, but anticipated in Flood deposits (Studies in Flood Geology,
pg 209-211).
Plant fossils of species believed to be extinct
are also frequently found encased in sedimentary rock supposedly hundreds of
millions of years after they were gone.
For example, fossils representing strictly Carboniferous floras have
been consistently found in the Jurassic throughout
Despite the obvious interpretation of the fossil
evidence for a global flood, the abundant fossil anomalies that absolutely
shatter the scientists’ own evolutionary scheme are simply disregarded as
“misplaced fossils.” That is
straightforward enough. According to
evolutionists, this evidence is simply not what it appears to be. They don’t seem to need to consider evidence outside
their expectations. It is physically
unexplainable to assert that any plant (or animal) material could burrow
through all that strata and delicately deposited itself, intact, within the
confines of the hard, sedimentary Cambrian rock. Without the use of a flood.
The fossil record unambiguously testifies
against the evolution of plants. It
shows us that there is no shadow of transitions to help plot the progress of
plants. The sudden worldwide appearance
of these fully formed plants in the strata could not be the result of gradual
evolution. The great range of fossilized
plants allows ample opportunity to capture these supposed transitions as well,
but only they are omitted. The Flood account is the only logical and
scientifically plausible explanation for the evidence revealed in the fossil
record. As the water overwhelmed the
land, the plants were either immediately covered or catastrophically torn up,
and floated in the waters, until they randomly sank and were buried by
sediments. A phenomenon that we have
witnessed numerous times with our own eyes.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Arthropods
There is another class of life that should be
addressed. In addition to the thousands
of plant and vertebrate animal species, arthropods form another group that is
not typically included in evolutionary discussions. Specifically, insects and arachnids
constitute an incredible variety in form and function. It is baffling to imagine how evolutionary
processes through time, chance, and mutation, could so extravagantly produce
such intricate and varied life.
Evolutionists contend that the fossil record
does not adequately preserve the history of insects, by asserting that
naturally they evolved, but that insects in general are not well
represented. This premise is false. What is not represented is any trace of
evolutionary development. Insects, however, are actually well represented in
the fossil record, and reflect what we see today from the very beginning. They have always been a versatile and highly
specialized group of creatures, fully developed from the start.
The reader will not be surprised that there is
no evolutionary history for insects in the fossil record. These marvels of miniature appear to be every
bit as developed in the past as they are today.
The fossils also demonstrate that there were even more variations in the
past, and that many insects were capable of becoming enormous. The cockroach and the dragonfly are some of the
lowest terrestrial creatures recorded in the “400 million year old”
Pennsylvanian rock. Supposedly these first
creatures to think of flying apparently did it with no transition from life in
the water to life in the air, though not a single preceding marine creature
resembles them. Evolutionists can only point
to trilobites for a possible insect ancestor, but there is no development
between them and the wide radiation of terrestrial arthropod and flying insect
species that appear suddenly and fully formed.
Although
insects and arachnids are supposed to be very ancient, looking at the fossilized
remains of gnats, or dragonflies, or spiders clearly traced in sediment, or a
mosquito or ant encased in amber, will reveal that they are indistinguishable
from today’s forms. People who know
about such things, fossil hunters and paleontologists, are amazed by insects’
lack of developmental history. The
Nature Company Guides Rocks and Fossils politely makes this concession on
page 111, although the book takes a strictly evolutionary assumption:
Insects, the most diverse group of metazoans in the world today, do not
have a fossil record to match their current massive diversity.
We don’t know how they evolved because the
fossils only record their final body form.
Insects and spiders would have been just as vital to a developing
ecological system as they are today. In
fact, they fit so perfectly into the ecological world, that we cannot imagine
life getting very far without them. Ants, bees, flies, spiders,
termites and beetles, are all indispensable to our ecosystem, and provide
essential services in the perpetuation of a balance of life. In a time of delicately evolving systems, insects
would have been necessary to carry out the same functions they do today. They breakdown dead matter, spread seeds, aerate
the soil, pollinate, and serve various other functions.
What evolutionary providence produced such a
wide variety of beneficial and marvelous creatures at the sacrifice of long
life? One would think that the brevity
of insect life would be contrary evidence of upward evolutionary success, but,
luckily, “evolution” very prudently provided these varied creatures with the
ability to produce vast numbers of offspring, and a multitude of cleaver
devices for survival.
Examining the variety of insect and spider life
and their methods of survival is like perusing a spy gadget convention. Every part of their micro-bodies is designed
for their specific function in the world.
There is incredible engineering involved in the different forms of flight,
the special hairs on their feet for suction, the unique method of respiration,
wide variety of eyes, the different eating, stinging, biting, and poisoning
methods, mechanisms for defense, for attack, for hunting, for hiding, for
reproduction and providing for offspring, and incomprehensively so much more,
testifying of their perfect design. But,
even in the lowest fossil finds of insects, there is no trace of development
from the sea to the land, where arthropods enjoy the most numerous and diverse classification
of animals on the planet.
There would have been extraordinary changes
necessary to get arthropods out of the water and into their land forms. In order to keep from drying out, major
metabolic transformations would be necessary.
They would also need to get rid of their gills and obtain a trachea,
develop those cool compound eyes, and turn their shells into wings, along with
innumerable other adjustments.
How did the beetle and other unrelated insects
come up with juvenile tissue disintegration when they decided to try complete
metamorphosis? Although dragonflies and cockroaches undergo
incomplete metamorphosis, ants, mayflies and beetles, which pass through
complete metamorphosis, supposedly soon followed. The fact that the fossil record is silent on the
development of hundreds of thousands of basic terrestrial arthropod species,
tells us that these dramatic events could not have occurred.
Already discussed are the perilous metamorphosis
of the butterfly, and the explosive arsenal of the bombardier beetle, which
both, of course appear fully formed in the fossil record. Another great example is the amazing
arachnid. These creatures have been located as “early” as
400 million years ago according to the evolutionist interpretation of their
fossils. Spiders are best known for the
silk they spin to form webs and traps.
In fact, all spiders catch pray, and all, but a few exceptions, do it
with their silk.
It is easy, due to the spider’s commonplace
existence, for us to overlook the uniqueness of this specialized trait. These “early” fossilized
spiders
and spider parts have been found with fully developed spinnerets, demonstrating
that spiders have always spun silk. As
mentioned in the Biology section, every spider species has a unique web design or
use for its silk, which every species knows intuitively. One wonders, at this
early stage in terrestrial evolution before most flying insects were supposed
to evolve, what the spider caught in his marvelous web.
Setting aside the ingenuity of the spider’s
engineering marvels in web design, the silk itself is remarkable. Three to five types of silk are made by each
spider, with chemicals and proteins produced by the body. This liquid is perfectly balanced and blended
with enzymes, then shot out and spun through uniquely engineered spinnerets,
using special hooks on the hind legs to lay down the line. This silk is the strongest natural or manmade
substance on earth. It is stronger than
steel, completely flexible, non-water soluble, and extremely elastic. Scientists think that if they could figure out
how to reproduce it, they could revolutionize the world. But they can’t. How did
the little spider do it without any intent to do it at all? If intelligent humans can possess the
substance they are trying to imitate, with all the resources in the world at
hand, and still not be able to do it, how could the spider happen upon it
through sheer accident and unerring steps in mutation? We can’t know this because the fossil record
is silent.
The insect world is full of these clever little
devices and surprising little quirks that “Mother Nature” could never produce
through thousands of accidental mutations for each. Humans can’t even produce such prototypes on
purpose, with the whole blueprint before us.
In light of no other plausible explanation, and the absence of any
fossil pathways, Creation scientists see the uniqueness of bug life as clear
evidence for Special Creation.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Birds
There are still many rumors dispensed to the
public about fossils that “irrefutably” prove evolution, but the myth of
transitional species has been built on a few promising finds that have since
been reclassified, or are currently in dispute even within the evolutionary
community. The favorite example of a
“transitional species” is Archaeopteryx, touted as a genuine intermediate between
dinosaurs and birds, and considered by many uninformed as an undeniable fact of
evolution. This bird was found fully
formed and placed by evolutionists in the Jurassic system, right when dinosaurs
were supposedly at their peak, which is considered still over “a hundred
million” years before “modern” birds.
Unfortunately for evolutionists, even their own
scientists recognize that Archaeopteryx is indeed a genuine bird. Many try to discredit Archaeopteryx as a true
bird with careful wording, by referring to it as a dinosaur with feathers. However, even the illustrations evolutionists
provide for Archaeopteryx betray this characterization, and show that they were
fully functional birds. The fossil casts of
Archaeopteryx clearly reveal even to the untrained eye the avian features of
thin, porous bones, perching feet with claws, avian pelvis, the small bird
skull with large eyes, a wishbone (only found in birds) and most significantly,
the gracile wing bones with all the perfectly formed feathers
meticulously captured in the impression. The whole of the argument that this species is
an intermediate between dinosaurs and birds rests on a few features that are
not common to birds today.
One feature is an apparent lack of a full keel,
or breast bone, though to what degree this cartilage may be preserved
protruding from the front of the bird may be a good question. Whether or not there was a full keel is
unimportant since the other features prove that the bird could fly, and was
likely a good glider.
Of the other unique features, there is also the presence
of small teeth in the beak, a bony tail where the feathers attach rather than
the typical nub, and the presence of small claws at the ends of the wing bones
(like the ostrich, swan and ibis).
Evolutionists have tried to make these features dinosaur-like in order
to classify Archaeopteryx as a primitive transition, but in every way, this
crow size species is unquestionably (even to evolutionists) a bird that could
fly.
The few unique features found on Archaeopteryx
are not completely bizarre, as parallel features have been found individually, though rarely,
in other birds. The tendency is to see Archaeopteryx’s unique
features as inferior to the typical bird today to allow for a purely
transitional status. But what if these
features were designed for a specific purpose?
Then, they would not be seen as intermediate features, but as
appropriate equipment for that species’ lifestyle.
What if, for example, Archaeopteryx was
predatory, and needed these features to glide from tree to tree and hunt for
the large insects that were more prevalent during that time? One could see how useful its apparent lack of
a full breast keel would aid in maneuverability in the trees. Its long neck and a beak lined with small,
sharp teeth would be perfect for digging into borough holes and gripping large
beetles and larvae. The stiff, bony tail
would help give the predator balance and leverage, the way the woodpecker’s
ultra stiff tail feathers do. The claws
on the ends of the wings would be necessary for gripping the tree trunk while
digging into the bark crevices.
Perhaps, altogether, Archaeopteryx was simply a
highly specialized bird in a time of a greater diversity of life. When we look at these features in such a
light, we can see that our presumptions can close our minds to other
possibilities. It is clear that we do
not know enough to be able to categorize a creature simply by examining its
bones, frozen in a moment like a death mask.
By emphasizing the small differences between living bird forms and
Archaeopteryx, evolutionists are trying to bolster a theory that could never
hope to provide real examples of transitions for birds because birds are so
incredibly unique.
When we use our own minds, and our own
observations, the truth is plain.
Archaeopteryx is fully a bird and it does not demonstrate a true
intermediate form. It absolutely
provides no information about the process a dinosaur would have to go through
to develop a bird’s highly specialized features with functional wings and precisely
designed feathers. But this is not the
real problem that Archaeopteryx presents for evolutionists. It gets worse.
Since the evolution of birds from dinosaurs
would have undoubtedly taken millions of years, and countless generations of
participants, there should be numerous fossils to document this incredible
transition. There is no corroborating
fossil evidence to reveal how dinosaurs could have developed the most
remarkable bird features, such as porous bones, functional wings or the
precision of feathers from their non-follicle skin.
There is a great deal of evidence, however, of
the artistic license abused by some paleontologists who frequently impose bird
features on dinosaur fossils in presenting their interpretation to the
unknowing public. The Natural History
Museum Book of Dinosaurs is an evolution-based book geared toward
teenagers. The authors admit to this
“enhancement of the truth” phenomenon on pages 71-72:
Even more controversial is the question of
whether some dinosaurs had feathers.
This remarkable idea was first suggested in the late 1970’s by the
Russian Scientist S.M. Kurzanov when studying the small meat-eating Avimimus. He thought that the upper arm bones of this
dinosaur showed feather attachment points similar to those that can be seen in
modern birds. If Avimimus had this feature, Kurzanov reasoned, other related
meat-eaters probably had it too. The
evidence is disputed, but has been taken up enthusiastically by a number of
dinosaur experts and illustrators.
Why Feathers?
It is generally agreed that birds evolved from small meat-eating
theropod dinosaurs. Obviously these
dinosaurs did not sprout feathers overnight and become birds, so there must
have been a long period when some of the small theropods were experimenting
with feathers, which are only a different version of scales (what on earth does that mean?) . . .
It is quite legitimate, say the feather enthusiasts, to illustrate such
dinosaurs with downy feathers, neck plumage, arm and even tail feathers. The American artist Gregory Paul draws all
his small theropods with feathers as a matter
of principle, ranging from fluffy baby Velociraptor to adults with virtual Indian Chief
head-dresses. Such ideas may not be
generally accepted, but they show how
far some dinosaur experts are prepared to go in stressing the direct
link between dinosaurs and birds. (author emphasis and parenthesis added)
This quote plainly acknowledges the
lack of compelling evidence for dinosaur to bird evolution, and reveals the
ease with which distortions of the truth can be inserted into the scientific
realm unchecked. Thousands of Creation
scientists are battling for opportunities to be published in established
scientific journals with legitimate and authenticated documented evidence. All the while they are being shut out, the
unsubstantiated assumptions about how evolution must have happened are
given legitimacy in forums void of true scientific scrutiny, and dispensed to a
trusting public.
And while this candid insight into the pliability
of science is rarely exposed (and clearly winked at), it shows that although
many evolutionists recognize there is a lack of authentic evidence for such
assumptions, it is to their advantage not to point it out. After all, if another scientist or
publication chooses to disseminate the fabricated information as if it were a
fact, why should it be widely exposed when it aids public perception? This use of fiction to substantiate evolution
reflects a level of scientific incompetence.
Since there is no other way for a dinosaur to become a bird,
evolutionists assume these imagined transitions have to be true, even if there
is no evidence to demonstrate them.
But the most detrimental fact against
the evolution of the bird from dinosaurs is that the evolutionists’ own
evidence and timeline invalidate every one of their proposed
transitional candidates. Each dinosaur
fossil that is supposed to be a bird in transition has been dated by
evolutionists as living after the remarkable
Archaeopteryx. Do you need to read that
again? To rephrase: the evolutionists
have NO dino-bird transition candidates that precede
Archaeopteryx. They all come after
Archaeopteryx by more than “25 million years.” Do you
understand that? Archaeopteryx, a fully
formed and functional bird, would come before the oldest dinosaur that might
hint at the slightest development of bird features.
In other words, a real bird came before any of
the transitions to a bird even started.
Funny how no one seems to think this is a problem. Paleontologists keep insisting that this
dinosaur or that one has “feather- like filaments,” as if these specimens could
have evolved into birds, when according to their timeline, Archaeopteryx would
have already been flying around. What
good is a dinosaur with mysterious filaments 25 million years after
birds already existed? Perhaps
Archaeopteryx was a bird born out of its time—an anomaly that evolutionists
disregard.
We shouldn’t even be entertaining bird
evolution, but since evolutionists never make a point of this great
contradiction, and one can only discover it if one happens to pay attention to
what geologic age the specimens are dated, and knows the order of the
geological ages. Obviously
paleontologists could find no way around the time placement of the findings
without corrupting the integrity of the evolution paradigm, or they certainly
would have done it.
This incredible “first” bird supposedly appears
in the fossil record at a remarkable time.
To reiterate, the numerous Archaeopteryx fossil finds have been placed in
the late Jurassic, which is considered the dinosaur era, more than 100 million
years prior to when “modern” birds are supposed to have first appeared. But all the supposed transitional candidates
are dated, by the evolutionists’ own system, millions of years after
Arcaeopteryx in the early to late Cretaceous.
In addition to appearing too late to evolve into
the 2 foot Archaeopteryx, the proposed candidates themselves further degrade a
feasible pathway: Avimimus at 5 feet, Sinornithosaurus at 3 ½ feet, and
Compsognathus at 6 feet. This last gawky
candidate is one of the “earliest,” but since it has been dated contemporaneous
with Archaeopteryx, it logically has no evolutionary relationship. Additionally, there are no wing features
present in any of them, and the insistence that many of these dinosaurs
possessed feather-like-filaments, whether or not the interpretation is correct,
still cannot be applied backward, since the bird already existed.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Evolutionists persist with this façade of
dinosaur to bird evolution to the point of ludicrousy. In May 2002, the great unveiling at Chicago’s
Field Museum of a complete 40 foot tall Tyrannosaurus Rex
called “Sue,” revived this preposterous notion with fervor. What ever drugs the public must be on to
accept this bizarre link to birds, ought to be pretty good because such a stir
should not be created by a gigantic animal that supposedly lived in the late
Cretaceous, also millions of years after
Archaeopteryx. Rather than risk
unveiling the impossibility of such transitions by their own assessments,
evolutionists confidently imply through their careful wording and innuendo that
they have a handle on the dino-bird transformation.
Such an early appearance of Archaeopteryx,
however, requires special handling. The
2002 reprinting of A Guide to Dinosaurs, (originally published by the
same people who produced The Nature Company Guides Rocks and Fossils),
crafts the preferred slant in order to create transitional implications. Page 130 on Archaeopteryx reads:
. . . because Archaeopteryx displays an
unambiguous mix of characters from the two linked groups of animals—the birds
and the dinosaurs—it is a classic and rare example of an organism on an
evolutionary pathway between the two.
Archaeopteryx was a small, birdlike
dinosaur, about the size of a present-day crow. Its skeleton is similar to that of some
theropod dinosaurs . . . The most strikingly bird-like feature
of the Archaeopteryx is the feathers.
Not only did this dinosaur clearly have feathers, but these were
arranged in exactly the same pattern as feathers on wings of modern birds. (hello, it is a bird)
Archaeopteryx
played a pivotal role in the acceptance of evolution as a mainstream scientific
theory. When Charles Darwin published On
the Origin of Species in 1859, a perceived weakness in his argument
was the lack of intermediate animals in the fossil record. If animals and plants have been changing from
one form to another through time, as evolution suggests, then at least there
should be some fossils of organisms intermediate in structure between the two
groups. The first skeleton of
Archaeopteryx was found two years after the publication of Darwin’s theory and,
as predicted, it displayed a mix of bird and dinosaur features. Clearly, evolutionists argued, it was an
intermediate form between the two groups . . . We now recognize that its
closest relatives are some theropod dinosaurs, such as the dromaeosaurs (6
feet, and from the late Cretaceous) and the oviraptors (10 feet, and from the late Cretaceous). In fact, the skeleton of Archaeopteryx is so
theropod-like that one specimen found without feathers was for many years
mistakenly identified as the small theropod Compsognathus. (how did they confuse a 2 foot
bird with wing structures and this 6 foot monstrosity, without any wing structure?)
(Parenthesis and emphasis added)
What is most astonishing is that the very information that
discredits these implied transitions is found right inside this same book. All the dates, sizes and illustrations
necessary to disprove the evolution from the dinosaur to Archaeopteryx are
provided under the description of each. It is no wonder the general public has given
up on discerning science with their own minds, when the scientific community
engages in such double speak, and misleading pronouncements. Evolutionists are careful to maintain a link
between dinosaurs and the “modern” birds placed by evolutionists in the
Quaternary, and are satisfied to imply the legitimacy of dinosaur links from
the late Cretaceous.
This, however completely skips over evolution’s
darling, Archaeopteryx—unless evolutionists would like us to believe that
Archaeopteryx was only a warm-up, and bird evolution senselessly had to start
over again. They must not want us to be
troubled by its untimely existence outside of the transitional chain. But since in their minds Archaeopteryx is so
handy for showing dinosaur features (limited as they are), if it is left out
entirely, there would be nothing to help bridge the gap between
birds and dinosaurs. Never mind that it
lived before the transitions that should have led to it.
There is even more devastating news for those
who would like to forget the “anomaly” of Archaeopteryx’s early appearance. One recent fossil,
Gansus, was found in the Xiagou formation in China, and is characterized as a
fully modern bird. The soft tissue
preservation was so exquisite that it revealed webbed feet, and appeared to be
some type of loon. Worst of all, evolutionists
were forced to date the fossil at 100 mya. That puts this highly specialized, fully
formed bird at the same age in the cretaceous as many of the supposed dinosaur
transitional candidates. How do they
manage to persist in this theory?
Evolutionists are quite innovative in the
perception they feed the public. The
July 1998 National Geographic (vol. 194, no 1) takes up this matter in
an article beginning on page 74, entitled “Dinosaurs Take Wing, The Origin of
Birds.” This pursuit takes 25 pages of
discussion and illustrations to establish the evolutionists’ claim, and only
one comment to note that the evidence does not actually substantiate it. The author states on page 95:
These skeptics of the bird-dinosaur theory
ask: If birds came from dinosaurs, why
can’t the paleontologists find missing links of an appropriate age? The majority of dinosaurs considered most
bird-like are younger than Archaeopteryx.
The pattern of deliberate
misconception about dinosaur to bird evolution is long, and unsubstantiated,
and getting somewhat tiring for scientists as well. Very little excitement has been generated by
the newest “transitional” find in China, dubbed Microraptor gui. This fossil
was found by Xu Xing, a
paleontologist at the Institute of Vertebrate Paleontology and
Paleoanthropology in Beijing, China, and it is believed to be a 3 foot dinosaur with
feathers on both the arms and the legs. However, few regard whatever it is as a
logical transitional species. They may
not be sure as of yet what to make of it.
Why? Despite the interesting
prospects, it does not really further their quest for a dino-bird transition as
it is again in the wrong time period, and far less developed than
Archaeopteryx. According to the January
2003 National Geographic online news article:
Although the M. gui fossils are about 25 million
years younger than Archaeopteryx,
the four-winged dinosaur is a more primitive form derived from a very early
evolutionary branch of dromaeosaurs
Although the fact of its improper
placement as a transitional species is more upfront, the obscure wording is
evidently an attempt to disguise this detriment. There is great confusion about what to do
with a poorly developed “transitional” fossil that places 25 million years after the fully functional bird
Archaeopteryx,
so they speculate that it is a branch off of the same ancestor that did develop into Archaeopteryx. An ancestor, of course, that they haven’t
found. There are many theories about
what this fossil could mean, and having not settled on one, silence steals the
excitement.
The evolutionary community makes plain how
dogmatic they are about this assumption of dino to bird evolution. We must assume that it is the undeniable
uniqueness of birds that compels evolutionists to infuse our perceptions with
this bizarre origin in these large, stout, scaled creatures. After all, if you can’t explain birds through
evolution, then evolution cannot be good for much, (the fossils don’t even
reveal how flying reptiles evolved). Despite
the lack of evidence within their own dating system, evolutionists insist to the
public that these remarkable things are true.
Their own words demonstrate that they recognize all the problems with
time, transitions, the necessary developmental stages, the lack of evidence,
and the willingness of the trusted “experts” to fudge it all. This is the science that fools the world.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
The Descent of Man
Though there are several other examples of
fossils of all types, which have been re-evaluated since their initial
discoveries, of the most interest to us are the remains of man, and the issue
of the missing links. The truth is, they
are still missing. In recent days, much
excitement has been generated about the “new missing link” in our supposed
ancestral chain. As you will soon see,
there has been no more ridiculous claim in the last few decades, and serves to
reveal the desperate sense in the evolutionary community that the public
appearance of continued progress must be maintained in the search for evidence
of primate to human transitions.
Regardless of the fancy TV specials and news reports, the “new primate”
Ida, or Darwinius massilae, falls so far short of what someone would hope for,
that one may feasibly conclude that the assertion is a deliberate propaganda
manipulation.
This find, by their own account, is assumed to be
47 million years old, has a long tail, and is
about the size of a small cat. We are
not looking for a 47 million year old link because paleontologists place the
first presumed step from primate toward Homo sapiens at about the time of their
famous “Lucy” no more than 4 million years ago—more than a 43 million year gap
on their own timeline. And “Lucy”
doesn’t have a tail. The new find is
clearly along the lines of a monkey or lemur, rather than the presumed
ancestral ape, as it has a long tail, and is quite small. In fact, paleontologists certainly are not
saying it is much more than a distant, pre-link, long before Lucy. However, one cannot imagine how the worldwide
hype promoted by this find could help the ape to man assumption in any way
other than to stir up the stagnant primordial pond to see if people will still
drink from it.
The true
burden of evolution on paleontologists is not the lack of monkeys in the
closet, but the utter lack of links from monkey to man. There is no skeleton, or even partial
skeleton, in the evolutionist’s closet to fill the gap between the 3½ foot ape
creature Australopithecus aferensis (a.k.a Lucy), and Homo sapiens. There is no evidence in
between the two that even evolutionists agree is a link. It isn’t for trying, either, as numerous
grand conclusions over the years have been drawn from some paltry bone
fragments, not even enough to outline the specimens’ major features. The entire support for primate to man
evolution is not based on evidence, but the presumption that it had to have happened because evolutionists
recognize no other viable option for how man came to be. Not only is there no evidence that all
paleontological evolutionists agree helps fill this great gap, but the
evidence that is offered as scaffolding for the unilateral
endorsement of this “history” is a distorted narrative about finds that few
paleontologists have the courage to publically correct.
A
History of Paleontological Mysticism
Such known distortions are permitted “for the
better good” of the theory, rather than draw attention to the severe lack, and
great fanfare, as in the recent find, is made of finds that are impotent in
transitional value, the way a magician redirects the eyes with an empty
flourish. It is common for whole
renditions of a creature to be divined from two or three teeth or bones, as in
the case of Nebraska
man. This entire family of “primitive
ape-man” was constructed from what was later determined to be a single extinct
pig’s tooth. The species Ramapithecus was
constructed, and then declared (later retracted) to be a perfect transitional
hominid based on only a few teeth and jawbone
fragments (most agree from an extinct ape). The scientific community is so eager
for confirmation, they can easily fall prey to bad science. In fact, Piltdown man
was an outright hoax, and a bad one at that. However, in such a receptive scientific
atmosphere, the artificial staining on the human skull
combined with an ape jaw, and filed teeth were not discovered for decades.
Evolutionists take great license in “drawing” their
conclusions (literally), and it is easy to conceal the true sparseness of the
evidence that these assertions are based on.
Surprisingly, all of human evolution is based on similarly rash
conclusions. When we examine the
evidence offered by evolutionists to bridge human ascent from apes, it
essentially breaks down into two distinct categories. The remains are either clearly apes, or
clearly humans. The transitional group
called Homo erectus is intended to exhibit the median of ape features and human
features. In reality, though, even
evolutionists cannot discern a true transitional example among the various bone
fragments, and are just as confused about the puzzle concerning which artifacts
reflect what trends.
The most questionable finds categorized as Homo erectus were
the first finds. About the turn of the
20th century, the scientific community was under a lot of pressure
to produce fossil evidence to support Darwinian evolution. Eugene Dubois heard about some possible
hominid discoveries, and traveled to
Dubois’ search yielded an unimpressive hodgepodge of bones
and fragments, most of which were not pertinent to his endeavor. In 1891 he found a skull cap that at first he
thought might belong to a chimpanzee, and some teeth that many conclude could
belong to an orangutan. The next year, and about 50 feet away, a fully modern human
femur was found. Dubois decided
that they all belonged to the same creature, and therefore behold, an ape-man,
or what we call, Java Man. Yes, this
grand example of paleontological detective work is still on the books, even
though his conclusion was initially rejected by the majority of his scientific
evolutionary peers.
Further evidence to discredit this conclusion was that fully modern human remains were found in the same level and
region not only by other researchers, but at least two were found by Dubois
himself. The subjectivity of the
evidence is malleable in the evolutionists’ hands. The skull cap fragment does not offer
sufficient evidence to conclusively determine its owner, which leaves its
significance open to interpretation.
Many argue it could be from either an ape or an archaic human. However, the femur is admittedly modern, and
found far away from the skull cap.
Combined with the other evidence of modern human remains guarantees that
Java Man cannot be taken legitimately as a transitional hominid species. After toying with several possible ages for
the finds, Dubois concluded that Java Man was about 700,000 years old.
Most Paleontologists view Java Man with caution, and regard
the femur as likely from a modern man.
Its fame would have easily died if any more encouraging evidence had
emerged. By the 1930’s only the
“Piltdownman” served to keep the evolutionary concept alive, as the 1912
“discovery” had yet to be revealed as a hoax.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
In the vacuum of true transitional finds, Peking Man was
discovered in
These fossils were studied for the most part by Davidson
Black and Franz Weidenreich. They
concluded that the skull fragments resembled the skull cap of Java Man since
they appeared to have a fairly large brain capacity (which they estimated
between 915 and 1030 cc), yet they were thicker than “modern” humans. However, no post-cranial skeletal fossils
were found to accompany any of the skull fragments. Despite possessing even less bipedal
corroboration, Peking Man was made possible on the foundation of the tentative
persistence of Java Man.
Regardless of the conclusions one would like to draw about
these pieced-together fragments of Peking Man, there is was no associated
skeletal evidence to demonstrate either imperative assertion for
evolution. If the skulls are interpreted
as ape-like, then there is no skeletal information to show that the owners have
any transitional hominid features. If
the skulls are interpreted as archaic humans, there is no evidence to suggest
that they are not fully human. The
evidence is too limited to draw any defensible conclusions about their
status. More alarmingly, all of these bone fragments were lost in 1941 when
researchers were attempting to get them out of the country for safety during
the war. Based on the fragments, models
were made that filled in the missing cranial features. While casts were made of the original
fragments, the fossils themselves have not been available for examination since
1941.
This fact alone is disconcerting. We must therefore rely on the original
researcher’s assessments (biased or not) of how the fragments fit together, the
conceptualization of the features that were filled in, and the estimation of
the brain capacity. But rather than
dismissing the finds on this lack of evidence (which would be reasonable), an
honest assessment of what is presented is sufficient justification. No evidence has been offered (that even
paleontologists recognize) to establish whether the specimens were fully ape,
transitional hominids, or archaic yet fully humans. Nothing.
There are only incomplete craniums, in fragments without jaws, without
limbs, without skeletons.
Many credible scientists believe the entire spectrum of choices for
their identity.
Now, it is important to consider the other controversial
evidence. These
fragments were found with an array of tools ranging from stone to sophisticated
iron. Subsequent research at the site
uncovered an additional 100 “modern” animals, more tools, and 6 “modern” human
skulls. Many conclusions have
been drawn from this evidence. Most
agree that it reveals that true humans had been busy at that site, and the
animals and tools found in abundance there indicate activity that borderlines a
high level of industry.
Even if the models of the original fossils and their
assessments are accepted, there is nothing to substantiate the conclusions
drawn about Peking Man’s status as a transitional hominid. Without any post-cranial bones—no
body, no limbs—there is nothing for this “ape/man” species to stand on. And certainly no indication if and how
it stood. Peking Man was dated at
between 500,000-300,000 years old.
This is the extent of transitional evidence available to
fill the gap between Australopithecus apes and true humans. There are many more finds, (almost entirely
craniums and skull fragments, again, without post-cranial skeleton
finds) but a close examination reveals that even the best examples offered as
Homo erectus are likely just humans with “archaic” features. When we recognize how contrived the
depictions of Java and Peking Man are, this transitional category
disappears. It is only perpetuated by
paleontologists who group the distinctly human fossils into the Homo erectus
category with these more transitional images constructed from the Asian finds,
lending a false credibility to the transitional concept.
Here is a quick list of fossils tentatively listed in the
transitional Homo erectus category. Sangiran 17, or “Pithecanthropus VIII” is a cranium (minus
jaw) that was found on Java in 1969. It
has brow ridges, thick cranium, and flaring cheekbones, but the eyes, nose, and
shape of skull appear to be very human, with a brain size of 1000 cc—within the
human range. Since no other
evidence is available, this specimen could arguably be categorized as an
archaic human.
The fossil of Homo ergaster (KNM-ER 3733) is
another cranium that was found in 1975 in
One of the best examples is the rare find of an entire skeleton in 1984 in
Paleontologists cannot deny that this long-boned skeleton is
as advanced as a modern human, and yet classified as an extremely early Homo
erectus, it should be far less advanced than the much later Neanderthals. Yet the most defining feature of Neanderthal
is their stocky, seemingly archaic skeleton, with the barrel chest and robust
bones. Turkana Boy is supposedly an
older transitional ancestor of Neanderthal that preceded them by 1.5 million
years. This timeline would mean that
Neanderthal apparently lost the advanced progress of this supposed walking ape,
making the developmental relationship untenable and superfluous.
Such conflicts are common among hominid candidates, and the
irregular evidence of each site forces new compromises at every turn. Though Turkana Boy is the only Homo erectus
with any known associated post-cranial skeletal bones, these features are
exactly what make it most undoubtedly fully human. Here, the only opportunity for the category
Homo erectus to be validated actually muddies the assumption with a fully
modern physique. The hominid
classifications get more complicated.
Skeletal remains that have been classified as more modern
than Homo erectus are commonly associated with craniums that display bafflingly
analogous archaic features to Homo erectus.
The appearances of these features are utterly random, and defy
categorization, although they have received tentative labels from
paleontologists. Their placement in any
ancestral category is dictated by presumptions based on the associated
archeological site and level evidence, causing evolutionists to be cornered by
the timeline of their own assumptions.
Despite the obvious similarity in cranial features, Homo erectus and
archaic Homo sapiens are kept over a million years apart through evolutionary
preconception.
Here are some of the best examples. “Rhodesian Man,”
found in 1921 is a complete cranium, displaying the archaic Homo erectus
features and robustness, but it has been labeled Homo sapiens, or fully human,
with a large brain size of 1280 cc. Petralona 1 was
another cranium discovered in
Atapuerca 5 found in 1992 in
In
This chart is to help keep a perspective of the average
brain sizes in attempting to gage the likelihood of “hominid” relationships.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Chart of Hominid Brain Sizes
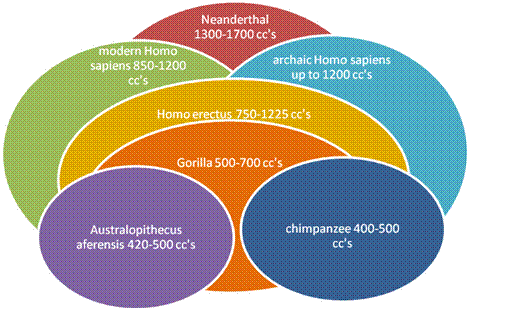
These brain sizes demonstrate that the supposed pre-human
remains from Homo erectus through “modern” are all within, and even exceeding
the human range. The wide 750-1225 cc
range allowed for Homo erectus demonstrates how ill-defined this fragmented
category is, which has permitted a wide spectrum dissimilar finds. There are no accepted transitional remains to
stand between true apes and the human range.
Now that the major finds have been discussed in terms of
their physical descriptions, this chart puts the evolutionist’s dilemma further
in perspective—according to the time they supposedly lived:
![]()
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
+ 3.5 m 3 m 2.5 m 2 m 1.5 m 1m
500 k 300k
100k 50k 25k present
![]()
Major Hominidae Finds
Australopithecus aferensis (Small ape) 3.5
mya +
Homo habilis (“advanced” ape) 1.9 mya
Australopithecus
boisei (Large ape) 1.7 mya
![]()
Homo erectus Pithecanthropus VIII 1.7 mya
![]() Very
early H.e. in Java
Very
early H.e. in Java
Homo
erectus “Turkana Boy” 1.6
mya
![]() Early,
gracile, 5’3 fully human skeleton
Early,
gracile, 5’3 fully human skeleton
Homo e. “Peking Man & Java Man” 700,000-300,000
ya
![]() Skull
caps and femurs of undetermined relationship
Skull
caps and femurs of undetermined relationship
Homo sapiens Petralona I
500,000-250,000 ya
![]() Homo erectus/ Neanderthal in early
Europe
Homo erectus/ Neanderthal in early
Europe
Homo sapiens Atapuerca 5
300,000 ya
![]() Archaic
semi-Neanderthal in
Archaic
semi-Neanderthal in
Homo sapiens “Swanscombe” 300,000
ya
Early archaic human skull in England
![]() Homo sapiens “Rhodesia Man”
200,000 ya
Homo sapiens “Rhodesia Man”
200,000 ya
Large brained advanced archaic
human
![]() Hs Neandethal Croatia 130,000
ya
Hs Neandethal Croatia 130,000
ya
![]() Partial
cranium; very early
Partial
cranium; very early
Homo sapiens “Omo I” Ethiopia 130,000
ya
![]() Early
fully modern skull/ part skeleton
Early
fully modern skull/ part skeleton
Homo sapiens, “Skhul” 1932 90,000
ya
![]() Early
true human skeleton in Israel
Early
true human skeleton in Israel
Homo sapiens, “Quafzeh IX” 1969 90,000
ya
![]() Early
true human skeleton in Israel
Early
true human skeleton in Israel
Hs Neanderthal (various) aprox.
70-35,000 ya
![]() Characteristics
vaguely grade between Homo erectus and modern humans
Characteristics
vaguely grade between Homo erectus and modern humans
Homo sapiens Cro-Magnon 35-10,000
ya
![]() Fully
modern humans
Fully
modern humans
Hs Neanderthal “Amud I” 28,000-6,000
ya !
![]() Controversial
late date for Israel, and uncharacteristically tall
Controversial
late date for Israel, and uncharacteristically tall
Archaic humans “Kow Swamp” 15,000
ya or less
Very recent though identical to Homo
erectus/ Neanderthal
Interpretation of Data
These examples are just an outline of the conflicting
conclusions that evolutionists are forced to draw based on the confines of
their necessary timelines. There are
many finds that prove that true humans and those labeled Neanderthal lived at
the same time, in the same regions.
Evolutionists are forced by their own assessments to acknowledge this
fact, which negates the basis for progressional assumptions. Paleontologists are frequently candid
concerning the subjectivity of their conclusions about the differentiation
between remains that are classified as transitional to Neanderthal, and those
labeled Neanderthal, and those viewed as early true humans with archaic
features.
If indeed even Homo erectus features cannot be definitely
distinguished from the much later Neanderthal, and Neanderthal grades into
later humans, which coexisted, then there is no real establishment of these
transitional categories. The locations
of the finds with their corresponding dates are not conducive to a logical
pattern. The Homo erectus “Turkana Boy” is over a million years older than Homo
sapiens “Atapuerca 5” which is just across the Mediterranean, but remarkably
similar in appearance. However, Turkana Boy is contemporaneous with the ape
Australopithecus boisei, and a million years older than Java Man and Peking
Man. Atupuerca 5 the Homo sapiens is therefore also contemporaneous with
Homo erectus.
Neanderthals have been limited to
between 70- 35,000 years ago as archaic humans, but Omo
I in Ethiopia is accepted as fully modern Homo sapiens despite the early date
of 130,000 years ago. Omo II, however admittedly resembles
archaic or Neanderthal humans, though the two were found close together.
Three last “transitional” species to cover are
Australopithecus robustus, Australopithecus boisei, and Homo habilis. These three are clearly apes. However, some of the skulls have been found
in such fragments that even evolutionists argue over the accuracy of the
identity and interpretation of these reconstructions, such as Homo habilis
1470. Skull 1470 caused another controversy
in that the original potassium argon test of the site
dated the find at about 220 million years old, which was clearly
unacceptable. Additional dates yielded
about 3 million years, which was closer, but it was
finally re-dated at the more marginally acceptable 1.9 million years old. Even after painful “readjusting” of the
radiometric dates, the final “accepted” date still strains the evolutionary
timeline, putting this supposed human link at the same time period as their
Australopithecus “ancestors.”
The fragmentary nature leaves a lot of room in these
specimens during reconstruction. While even evolutionists remain unsure as to
the extent and existence of these categories, all assessments and evidence
confirm their clearly ape features.
Though it is difficult to completely discern the features of some of the
fragmented fossils in the Homo habilis category, evolutionists are not even
sure which specimens belong there. The
absence of skeletal evidence for bipedalism in all instances, and contemporaneous
existence with a supposed descendent (Homo ergaster) eliminates any basis for
these three as transitional body types.
There is a complete lack of any transitional evidence to
demonstrate a tie between apes and humans.
The evidence used to represent Homo erectus offers nothing to
substantiate this supposedly ape-man category.
Java Man and Peking Man do not even meet paleontologists’ criteria for
demonstrating a bipedal transition. The
on-site evidence of modern humans further discredits the conclusion. Outside of these two, the rest of Homo
erectus is arguably comprised of humans with archaic features that grade both
ways into Neanderthal Homo sapiens and modern Homo sapiens. Once again, evolutionists are left with an
untenable progress from apes to man using just apes and just man. And they are quite aware of this
predicament.
When interviewed in a PBS documentary in 1990, a member of
the great paleo-anthropologist Leakey dynasty, Richard Leakey, expressed the
lack of hominid transitional evidence:
If pressed about man’s ancestry, I would have to
unequivocally say that all we have is a huge question mark. To date, there has been nothing found to
truthfully purport as a transitional species to man . . . If further pressed, I
would have to state that there is more evidence to suggest an abrupt arrival of
man rather than a gradual process of evolving.
One might say that God was the source of this abrupt
arrival. Unfortunately, although there
are no fossils that actually show a genuine transition, the famous names of all
these baseless links stand in for facts in the public mind. They have no idea how little evidence these
icons are fashioned from. Between apes and humans there reside only an
assortment of more apes and more humans.
There is no intermediary evidence of any kind
to flesh out the evolution of man from primates.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Here’s Lucy. . .
There is no hominid transitional specimen, but
evolutionists still put a great emphasis on what they believe is the jumping
off species of human evolution—a walking ape.
Although the greatest such specimen for paleontologists is the Australopithecus
afarensis called Lucy, the first famous Australopithecus called the “Taung
Child,” was found in what is now
The Taung skull was met by the scientific
community with mixed reviews. The
assessment of its cranial features was not so much scientific as it was
subjective. The general consensus was
that, although cute, it was very apelike, and didn’t demonstrate anything
concrete to establish its hominid status.
As an adult, however, aferensis is clearly apelike—even more apelike
than the modern bonobo chimp. Despite
scientists’ evolutionary assertion about the Taung fossil, what the skull
should have demonstrated went overlooked.
The skull was so well
preserved because it was found encased in limestone sediments, which even preserved the
mandible, and filled in the braincase.
What possible circumstances lead to this type of
fossil? Did the creature lose its head,
with the jaw, and fall into soft pool of limestone, which instantly preserved
it from predators? To add to the
mystery, eggshells were found encased with the cranium. This is a most unnatural environment. Rather
than evidence of evolution, this find is the type of strong evidence that
points to the Flood. Similarly absurd
evidence will be discussed further later.
Following the Taung child, interest in
Australopithecus propped a scientific eye open for similar finds. Fragments categorized as such have been found
from
Lucy was found in 1974 by Donald Johanson. She is a collection of fragmentary bones,
which, as previously mentioned, resembles a 3½ foot species of ape. Lucy was a good specimen for paleontologists
because finally the majority of the skeleton had been located in one place. At about 40%, almost every bone is
represented on at least one side. While
Lucy’s structure certainly resembles the human structure (as do most primates),
the need for a hominid transitional jumping off point has been a powerful
interpretational motivator.
Australopithecus aferensis in general is an ape
species that has been linked as an early human ancestor based entirely on the
assessment that it could possibly walk upright. Though the fossil evidence of this species is
fragmentary, afarensis clearly retained the arboreal features for tree
climbing. Here is a list of features
that have been noted by paleontologists: ankles geared for tree climbing as well as long
curved hands and feet, and (arguably) opposable toes, with no discernable foot
arch, allowing the foot to grasp.
Australopithecus aferensis also had a higher
scapula that brought the chest cavity up to balance toward the head for tree
climbing, and a more apelike femoral head angle and articulation with the
hip. These features indicate that the
body weight is centered more between the legs and the arms for knuckle walking,
as indicated by the proportionally longer arms and shorter legs more designed
for climbing. Australopithecus
aferensis’ femoral head does appear to be angled slightly toward the human
position for a primate, but it is still consistent with a bonobo chimp.
The human femoral head, however, extends up and
dramatically away from the main shaft of the femur to center the weight of the
body in a straight line over the hips.
The comparison is difficult, as even the variation among humans yields
notable differences. Primates in general
are very visually similar in the femoral head, and yet their range of motion is
markedly different. In addition to the
similarity between afarensis and chimps, the femur in aferensis appears to bow
outward slightly—at least in Lucy. Other
primates with femurs that are similarly bowed, like in gorillas and gibbons,
have highly opposable big toes.
The aferensis pelvis also appears more elongated,
like other apes, with an inadequate sacrum for muscle attachment indicating a
similarly insufficient development of the buttocks. Humans require a broad, stout, bowl shaped
pelvis to anchor the powerful buttocks muscles, and cradle the internal organs
in order to walk upright. This required
physiology is not prominent in afarensis.
Gorillas, actually, have a more bowl-like pelvis, yet they clearly do
not walk in the human manner. Since
aferensis emphasizes an apelike opposable toe verses an opposable thumb, this
demonstrates the dependence on climbing and grasping with the feet verses
walking and using the hands for tools—one of the most humanlike things we do.
The evidence for bipedal locomotion rested
initially on the assessment of the angle of articulation in the Lucy specimen’s
knee joint. Since some of the physiology
does suggest a form of bipedal locomotion, this should be allowed, but clearly
not in the human manner. The features
just mentioned are admittedly suited for life in the trees, and not for walking
upright more than occasionally, and likely with an accommodating stoop.
In fact, evolutionist anatomists Jack T Stern
and Randall Susman examined Lucy, and other afarensis specimens in great
detail. In their 1983 publication of
finds in the American Journal of Physical Anthropology, they noted on
every count that Lucy was suited for arboreal life (in the trees), and did not
exhibit a specific increase in bipedal features. Even now, many anthropologists agree, it is
not sufficient to simply establish the chimp’s occasional ability to stand
upright (as any ape can do today) to determine that Autralopithecus afarensis
was part of the human lineage.
After further studies on “Lucy,” one leading
anatomist, Dr. Charles Oxnard noted several inconsistencies with an ancestral
link and concluded in his 1984 article “The Order of Man” (New Haven: Yale
University Press):
. . .the
australopithecenes . . . are now irrevocably removed from a place in the
evolution of human bipedalism, possibly from a place in a group any closer to humans
than the African apes, and certainly from a place in the direct human lineage.
The arm proportions and other features of
australopithecine, like those of other apes for ground walking and tree
climbing, offer no reason to infer that the animal spent an unusually increased
amount of time on two feet in relation to other apes. But apparently the angle of the knee joint,
and the possibility that an animal could walk on two feet is enough to
establish the factuality of our primate ancestry for most evolutionists. Never mind that most apes, prairie dogs, and
even bears do quite well on two legs, and when the Gibbon goes to the ground,
it usually walks upright. None of these animals,
however, are on their way to becoming human, and their manner of locomotion is
distinctly non-human.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Although there is no compelling evidence that
apes adapted to this human-like method, evidence that contradicts evolutionary
assumptions is ignored. Sadly, a well known site in
There is nothing to distinguish these prints
from modern humans, and no evidence in them of the divergent great toe, in-turned foot, or
even of knuckle marks indicating an aid in balance on the wet surface, yet logically Australopithecus should
exhibit these traits in their footprints. Most anthropologists have concluded that
these creatures did not spend the majority of their time naturally on their
feet, and yet they are supposed to have walked through slippery mud without the
aid of their long arms for balance—and in the manner and stride of a
human. Moreover, the prints exhibit the
heal shape of natural bi-pedal walker, and most importantly, a distinct arch in
the footstep, which is absent in aferensis.
The straight path and forward pointing feet indicate an expert walker,
and if they were found in the sand at the beach, they would be identified
unquestionably as belonging to a human.
Evolutionists rely heavily on their license to
manipulate the evidence as a substitute for clear facts in order to convince
the public that the theory is secure.
This book has already discussed how the Discovery Channel and other
science-geared networks create entire programs expressly to solidify evolution
in the public mind. The new tool is
computer animation, which is effectively used to create images and scenarios
that realistically model nature programs.
Instead of real life, though, these shows depict a “pre-historic” earth
with “pre-historic” animals and hominids by imposing the evolutionary
interpretation on the evidence.
They offer few facts to support their interpretations, and
use imagination to flesh out the paltry bone and fossil fragments. Through the magic of computer animation,
every aspect of “pre-historic” earth leaps to life through complete renditions
of species and their behaviors. These
powerful images sear into the minds of the viewers as being scientifically
authentic. According to this media
blitz, evolution and the ascent of man from apes are factually secure. However, when we examine the actual
physical evidence of the human lineage offered by anthropologists, we see there
are still no human-like fossils that even evolutionists agree are
non-human.
The public has this concept that whole skeletons
of these graduated steps through the evolution of our species are mapped in
irrefutable detail. In reality, little more than a
fragmented cranium or skull cap or teeth are typically used to generate the
structure of an entire transitional specimen.
Museums take reproductions of these fragments, and create every missing
feature with resin, and then cover them with the skin and hair and
characteristics as they see fit. They are then posed to give the
impression that they are almost human—although not
too human. All these contrived details
reassure the public that human evolution is a fact, and they would never
suspect the paltry and disputed fossil evidence that is intended to support
it.
Under these conditions, it doesn’t take long for
questions to arise within scientific circles because of the flaws in these
available specimens. A study of
anthropological discussions of these specimens over the decades reveals a great
deal of dissention, reevaluation and readjustment. Of the popular “species” in our family tree,
none of them “stand” on their own two feet in the gap between primates and
humans. Even among accepted primate
specimens, such as Australopithecus, they are effectively disqualified from our
family tree. The occasional
contemporaneous appearance of “true humans” with their supposed ancestors
further discredits their evolutionary lineage.
Primates and humans co-exist today, yet there is no link between them,
just as reflected in the fossil record.
The difficulty with this muddled lineage was
expressed by evolutionist Stephen Jay Gould:
What has become our ladder if there are three
coexisting lineages of hominids (A. africansus, the robust australopithecines,
and H. habilis), none clearly derived from another? Moreover, none of the three display any evolutionary
trends during their tenure on earth: none of them become more brainier or more
erect as they approach the present day.
Despite the occasional claims of new hominid
finds, there is still nothing concrete to link apes to humans but implication
and imagination. It appears that the
trail of human lineage grows cold after one ape, and jumps straight to
humans. Even the locations of all the
supposed transitional hominid finds that we have discussed pose serious
logistical problems. They span from
One theory, perhaps recognizing this strain on
the parameters of human evolution, proposes an unlikely adjustment. That, inconceivably, separate groups of
humans could have evolved from apes through independent lineages in different
parts of the world. But this cumbersome
concept is an unnecessary adjustment since the specimens that represent this
progress are rationally viewed as humans, and in recognizing their equal
humanity, the ruse of evolution is dismantled.
Evolutionary scientists have a great deal of
difficulty filling in the family tree with the evidence of fossils. Although many insist that the evidence is
there, even evolutionists are revising previous assumptions. The Atlas of Life on Earth,
acknowledges this lack of evidence on page 312:
. . . the evidence for human evolution is
patchy. The early stages, especially,
are poorly known, and theories for the origins and subsequent dispersal of
anatomically-modern humans are among the most contentious of all scientific
arguments of any discipline.
So many of the early specimens included in
hominidae are recognized as dead ends that even this evolution based book
concedes the legitimacy of the transition dispute. It too confirms, despite the scientific
community’s fervent insistence on the factuality of human evolution, that there
is no evidence to fill in between the Australopithecine apes and humans. Page 318:
The human family tree appears rather sparse . .
. Because the evidence is so meager, any tree is, at best, a summary and many
of the connections are open to debate.
The main division is between the small-brained australopithecines and the
large-brained, fully bi-pedal Homo.
Once again, what is contended to be a fact has
no facts to support it. If indeed there
are no fossils to fill the gap, and Human evolution has not been
observed, then what could this “evolutionary fact” be standing on? The true fact is that even highly
regarded evolutionary anthropologists from around the world cannot agree on how
to classify these fragmentary bones vying for a place in our human
lineage. The problems with each are
apparent, and the overall consensus is that Australopithecine is no more than
an extinct ape.
If those searching for validating links must
concede this, then why should the world still hold onto them? Scientists are not even able to plot the
general evolution of simply primates from the fossil record, let
alone specifically humans. By
saturating the public with all these fossils and theories, there is a general
sense that if the link has not yet been found, it is only a matter of time. It is practically in hand because the shroud
of scientific implication has veiled the issue, obscuring the truth.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Neanderthals
The branch of the human family tree is so
fraught with contradiction that Neanderthal and other “archaic” hominid
categories, which were once offered as transitional ape-man ancestors, are now
simply considered humans by all experts.
Neanderthals are recognized
as living both contemporaneous and in the same region as “modern” humans, casting reasonable doubt on the
certainty of their developmental relationship.
Evolutionists have capitalized on the “primitive” appearance
of the skeletal remains of Neanderthal and other “archaic” humans to bolster
the concept of man’s ascent from apes.
Since there are no examples of true transitional species ancestral to
man, these fossils are vital for lending any credibility to the assertion. However, evolution is not the only possible
conclusion that can be drawn from this evidence, and there are many known
factors that would rationally account for these divergent characteristics. The buffet that is about to be offered is not
intended to give an absolute definition of the cause of these perceived
differences, but to show how many authentic explanations are even more
reasonable than the unsupported theory of evolution.
One common line of thought that can explain the
identified Neanderthal characteristics is location. Since so many of the people classified as
Neanderthals have been found in more northern and European areas, many
scientists agree that the Ice Age may have had a profound effect on their
skeletal formation. Those skeletons classified
as Neanderthals are typically shorter, and more robust in build, with
characteristically large brow ridges, a thicker cranium, and often have other
prominent bone growth. Surprisingly for
evolutionists, Neanderthals also commonly have a larger brain case than the
human range, which raises serious questions about their “primitive”
status.
The Ice Age could have influenced the stature of
these people in a couple of ways. One is
that the group of people who migrated north during the Ice Age would have
naturally been those best suited for the cold climate, and the people who
succeeded there would have had the necessary variations available to help them
adjust. Such variations would include
stouter bodies to reduce the exposure to cold, a barrel chest to facilitate an
efficient respiration, and robust jaws firm from chewing tough, dried meat like
those people presently found in the arctic regions.
Additionally, there is some indication in a few
of the remains that nutrition and disease may have played a part in bone growth
and formation. Rickets, for example, is a
condition that increases bone malformation, and limits normal growth. It is caused by a deficiency of vitamin D,
which most people derive from the Sun. The Ice Age climate would
greatly reduce exposure to the sun, and hinder vitamin D intake from other
sources. Arthritis has been detected in others.
Additionally, the group of people that ventured
into the colder regions was possibly a somewhat isolated group. The motivating factor behind entering the
cooler climate might be the desire for a lack of competition. In such a case, this group could have
perpetuated a similar genetic pool, with very little fresh genetic input from
people outside the group, leading to common group features. But since remains classified as fully Homo
sapiens have been found in abundance with Neanderthal, there is no evidence to
support isolation as the major genetic influence. This is a larger task for evolutionists to explain.
While the Ice Age could explain some of the
features, others may have a different origin.
One of the most familiar features attributed to this group, a thick,
protruding lower jaw and extended upper front teeth, has been used by
scientists in the past to point to an intermediate stage between apes and
humans. Though many Neanderthals do
appear to have very robust lower jaws, recent studies have called into question
the interpretation that the lower jaw on Neanderthals protruded out in its
stereotypical apelike appearance at all.
By re-examining the actual skeletal remains, at
least one expert, Creationist and orthodontic expert Jack Cuozzo, discussed an
observation concerning the cranial bones of Neanderthal in his book Buried
Alive. In order to allow for the
present interpretation that Neanderthal lower jaws protruded in an apelike
fashion, it was necessary to set the lower jaw out of its natural
position. In this position, not only
would the jaw be dislocated, but the teeth clearly did not meet in occlusion in
this position. In other words, the teeth
fit together in a natural chewing position only when the jaw was set correctly
in its place. This gave the skull a much
more familiar human appearance, and demonstrated the subtle influence of a
strictly evolutionary interpretation of evidence.
Cuozzo proposes an interesting possibility in
his book. Based on his cranial and
orthodontic measurements, he compared the wide range of patients he has
documented, with actual Neanderthal skulls.
He believes that the formations noted in his older adult patients
compares to the more prominent features in Neanderthals. He concludes that the brow ridges and the
front of the mouth continue to add bone and elongate with age, distorting these
features. Given this tendency, he
believes that Neanderthals could possibly be rare examples of people who lived
to a greater age than we observe today.
In reference to the Biblical account, the
descendants of Noah reportedly lived hundreds of years, progressively dying
younger and younger after the Flood. One
must concede that once the rest of the Bible has been confirmed, it is
reasonable to consider that the recorded phenomenon of long life could be
accompanied by this effect on cranial bone growth. Certainly, the evidence of healed fractures in Neanderthals supports a concept of long
life and the capacity to recover from ailments.
There is one last factor to keep in mind
concerning bone malformations—a condition that today we call acromegaly. It is caused in adults when their pituitary
gland becomes over stimulated, resulting in abnormal growth in just the hands,
feet, nose, jaw, and brow ridge. This
condition can lead to significant enlargement of these features over the years,
and is usually caused by a pituitary tumor.
Medically, an increase in this occurrence even outside of colder regions
is a foreseeable effect after the Flood.
As mentioned, one of the assumed atmospheric
changes as a result of the Flood was the collapse of the water canopy. This loss of protection would allow more of
the sun’s radiation through, resulting in not only increased genetic defects,
but tumor growth as well. Perhaps the
bone growth was a combination of the preceding theory of long life, and a
poorly regulated pituitary. Either way,
we shouldn’t be surprised if this condition, which exists now were captured,
though sporadically, in the bones of a post-catastrophic past.
While it is important to consider these
authentic influences on bone formation, the simplest, explanation is likely the
most rational. All of the people of the
past did not necessarily contribute significantly to the population of people
we have today. After the Flood, it would
be reasonable to assume that as people disbursed, they took their little packet
of genes with them. In the wide-open post-flood
world, a few genes can go a long way, or they can come to an abrupt end. As people spread through the new world,
familial and racial variations would naturally emerge which would account for
the unpredictable occurrence of these noted features.
What we see in the bones of the past is a
smattering of unique features, from a relatively small anthology of
remains. Although there are certainly
enough remains to establish the frequency of these anomalous characteristics,
these examples still defy pure categorization.
By all accounts, nearly every ancestor with “archaic” features shares
distinct cranial or skeletal features with both modern Homo sapiens and/or
supposedly more primitive ancestor, which makes specific categorical
designations unclear.
Naturally when evolutionists do not use carbon
14 dating because the samples are too old, they must rely solely on their
evolutionary estimations. The factors
used in these assessments are completely subjective, such as the primitiveness
of features and presumptions about human technological progress. But when the appearance of these
characteristics themselves cannot be contained within a timeframe or pattern,
then the system is again arbitrary.
Therefore, it is logical to conclude that rather
than indicating evolutionary transitions, the smattering of prominently “archaic” features
that appear in ancestors from
Geneticists have taken up the cause to help
evolutionists demonstrate just how primitive Neanderthals were in hopes,
assumedly, to shore up the evolutionary ancestral relationship. At least two studies were conducted on, if one
can believe it, mitochondrial DNA extracted from Neanderthal
remains; one in March 2000, and another in March 2004. It is a phenomenal mystery how one manages to
get a readable DNA sample from over 50,000 year old bones that should not even yield
measurable amounts of carbon 14, let alone genetic material, which should break down in about 10,000 years. Perhaps (dare one suggest) they are not that
old. Regardless, the DNA was compared to
living humans, and whatever specific markers were assessed fell, they
concluded, outside the range of human variation.
In other words, there were more differences
between Neanderthal and modern living humans than between the average two
modern humans today. This seemed to
bolster the concept that Neanderthals were not only primitive humans, but
perhaps not even part of modern human lineage after all. Perhaps a separate species. Wow.
Really? Unfortunately, though,
the same test was performed on the anatomically modern “Mungo Man” of
Perhaps science is getting a little carried away
with itself. Perhaps one should not
expect DNA from hundreds of generations ago to fall comparatively within the
range of those who carry on the stock today.
Perhaps a lot of DNA has been exchanged and dropped over the centuries,
not to mention pooled together. A guy
with strictly one kind of DNA might be a little different from his descendants
with lots of bits and pieces of many people’s DNA spliced in. Maybe one should rather ask why the DNA is
still viable if the people are so ancient.
Scientists are not able to demonstrate a logical
line of decent incorporating their transitional perceptions of “Neanderthals”
and other people with “archaic” features.
These remains can rationally be regarded as people exhibiting just
another facet of historical humanity.
These people may be distinguishable by certain prominent features, just
like any race today, but these features are based on judgments about relative
size and shape. Although certain
features today can be closely associated with a type of race of humans, not all
people in the world who exhibit one of these features is necessarily of that
race, and not all people of a race will exhibit those certain features. “Archaic” humans follow this same pattern,
blurring the lines of ancestral distinctions.
Even today rare individuals may exhibit a
feature within the Neanderthal range, but they will not be classified as
Neanderthal. Such distinction is only
made concerning these people from the past, and only for the most prominent
examples. More damning is that any
remains found with these features are subject to being labeled archaic, and any
remains with more gracile “modern” features are labeled modern, without any
corroborative evidence beyond assumption to show that these characterizations
actually follow a progressive pattern.
Meanwhile, for all the scientist knows, they all existed at about the
same time, each having these different characteristics, just as we see in the
world today.
The designation of Neanderthal as primitive is
also not supported by on site evidence, which suggests that Neanderthals had
the physical, mental and social capacities that all other humans share. Artifacts such as advanced tools and paintings
and evidence of social structure and long life demonstrate that the people classified as
Neanderthals were intelligent, and lived as any human would under their
circumstances. They evidently lived in
caves when weather necessitated, and out in dwellings when it permitted. Iron tools have even been found at the same site
level, some (as in the “London
Artefact”) exhibit very refined forging processes. Clearly, the initial interpretation that
Neanderthals were not as smart as modern humans, or lacked the capacity to be
creative, or the dexterity to make tools was wrong. All of these assumptions have been revised by
further finds. There is no
paleo-anthropological affirmation of a primitive, transitional human.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO
Cavemen and Ancient Civilizations
Scientists embellish this evolutionary concept
of “primitive man” with “cave man” imagery.
Even elementary school textbooks discuss the “stone age” with full color
pictures as if there is actually scientific evidence that humans evolved from
primates through a stage of diminished brain capacity. These can be powerful images, but the premise
is unfounded, and the interpretation of the evidence is without merit. It is true that there are stone tools, and
cave paintings, but this is the limit of the evidence.
To bolster the evolutionary premise,
evolutionists make the case that people who use stone tools and live in caves
are less intelligent, and therefore possess a more primitive capacity, but this
is simply an assumption. Anyone today who became
stranded in the wilderness would be forced to use stone tools and live in any
available cave against harsh weather conditions until better arrangements could
be made. Even Native Americans were
found using stone tools when Europeans arrived, yet their brain capacity was
unquestionably equal to the “advanced” Europeans.
Societies tend to develop in certain patterns,
regardless of the technological developments available. People who continue to migrate do not tend to
progress technologically as long as they are able to keep moving to find
adequate resources. Historically, people
usually begin to develop technologically when they settle down into cities, and
build permanent residents. Lacking these
technologies does not mean that people are at a more mentally primitive,
evolutionary stage.
When American pioneers crossed the wilderness, and arrived
in the west, they did not build large luxurious concrete apartments to live in,
like those that had existed for centuries in
We would expect to find the evidence of periods of basic
survival throughout the world. Every
society had to go through it, and the older societies, the ones who stayed
behind and experienced overall stability, like in
Metal and stone tools are not datable except by
their styles, so any such
artifacts would be placed in history according to preconceived historical
perceptions.
Surviving ceramics are often used this way for dating as well, but
evolutionary assumptions, not carbon 14 dating, influence how the oldest
fragments are interpreted. This is not
to say that more recent archeological assessments about a society cannot be
trusted, but that the presumption of evolution has the biggest influence on the
interpretations of the evidence when a date cannot be determined accurately
through written record or other falsifiable means.
Moreover, evolutionists believe that humans did
not begin practicing agriculture until just 10,000 years ago. This is a tremendous admission because it
means that scientists cannot find datable evidence left by humans of any
significance prior to a few thousand years ago.
If evolutionists cannot stretch their own timelines further back because
of the constraints of the evidence, then there really is no evidence
that humans are older than 10,000 years.
A non-agricultural people would consist of nomadic tribes. Cultures don’t build cities if they are hunting
and gathering because they keep moving. There
is no industry, because they have to move on.
All non-agriculturally based cultures produce as
artifacts are the stone tools and the animal skins, temporal building materials,
and likely some pottery. The time when
stone tools are created can’t be dated radiometrically, and the skins,
temporary building materials and pottery wouldn’t last that long. Evolutionists clearly have reached this
recent date because there is nothing more significant to work with. Yet if humans had been around for over
200,000, what were they doing? What took
them so long to get to
Whenever evolutionists announce a new find of
human artifacts or remains, their assessment always reinforces their timeline
for a long period of human civilization.
Such remains are dated to be 20, 50, or 100,000 years old, and carry the
convincing weight of scientific authority.
People don’t realize that these dates are not asserted through some
infallible radiometric process (which is actually quite fallible) since
volcanic ash is rarely available for testing near the site on the level of the
find. Dates concerning human remains or
artifacts are calibrated entirely on evolutionary assumptions. Scientists frequently banter about their
assessment of an artifact’s age, and often reassign ages when conflicts
arise. This evaluation process is not
verifiable outside of evolutionary presumptions.
As will be discussed further in the geology
section, only carbon
14 can test the age of organic material or artifacts such as human remains, or
cloth, wood, papyrus, and rope that might have been fashioned by a human. However, once again, carbon 14 tests are only
fairly reliable within a few thousand years of the death of the organic matter,
if contamination does not occur. Since
carbon 14 begins to degrade rapidly, we are able to measure for ourselves the
rate of decay in relation to the remaining stable carbon 12.
Since no one has obviously lived 5,760 years to
verify the accuracy of the presumed half-life of carbon 14, other confirmations
are required to verify it. We would not
be able to account for every possible factor that could influence these ratios,
and there is no way of knowing for sure what could have affected them over the
centuries. Artifacts assumed to be older
than the half-life of carbon 14 not only enter a slippery range for accuracy,
but the sample would be subject to even more external factors that cannot be
accounted for. Anything over 50,000
years should have no carbon 14 at all, despite the common claim of artifacts dated
nearly this old and older.
Anything older than the oldest historical records
would yield less than half of the presumed carbon 14, and the interpretation of
the age of the remaining amounts becomes a tenuously delicate process, which
would be unverifiable beyond these written records. Artifacts from civilizations presumed to be tens of
thousands of years older than the written record would be assessing such a
small ratio of carbon 14 that minor miscalculations lead to large errors in
age. Artifacts credited with being tens
of thousands of years old are relying on samples where only a few atoms of
carbon 14 either way will affect the calculated age by thousands of years.
This narrow thread of evidence allows the assessment of age
to be dictated primarily by evolutionary assumptions—an unscientific biased
preconception. Therefore,
every archeological or human artifact that is dated to be tens of thousands of
years older than confirmed written human history of less than 5,000 years is so
dated based on preconceived assumptions about the progress of civilization from
our supposed primate ancestors. No artifact
or human remains can be proven to be more than 5,000 years old outside of these
assumptions.
Carbon 14 is best at confirming writings to
double check timelines. The
Writings still remain the best evidence of
chronology since it can be confirmed on many levels, and assumed that the
original writers were not intending to play a hoax on future generations. Other cross confirmations of the Tel Rehov site
come from Egypt. Pharaoh Shoshenq I
invaded
In fact, despite longstanding skepticism of the
Bible’s trustworthiness, archeologists continue to confirm its historical
accuracy. Some of these artifacts are the Ras Shamara
tablets, the Tel Dan inscription, Hezekiah’s tunnel, the Moabite stone, and
many more. Histories and references from external sources
continue to emerge, and by referring to events or kings, these events can be synchronized. The questions about the chronology of human
civilizations according to the Hebrew Bible are not raised because of
definitive external writings that disagree, or claim to have occurred at a much
earlier date, but because of evolutionary assumptions imposed on the evidence
about when civilizations existed.
Even attempts to discredit prophetic writings,
such as Daniel, are refuted by a forensics style examination of the
language, which confirms the text’s date.
The Biblical book of Daniel makes such astonishingly accurate
prophecies of the events that would come hundreds of years later, that rather
than acknowledge the divine inspiration behind the prophecies, scholars have
tried to discredit the book’s attested date of c. 530 BC. Instead, in light of the specific prophecies
about the future of the Persian, Greek and Roman Empires, scholars claim that
the Book of Daniel was actually written c. 2nd century BC,
and not by a Jewish captive in Babylon.
This presumes that the entire book is a work of historical fantasy, and
not one of historical record and prophesy.
However, there are at least two major problems with this theory. Firstly, the writer’s specific references and
descriptions of cultural and political and historical details are perfectly accurate,
which would have required extensive and likely inconvenient research by a 2nd
century BC author just to perpetrate the fraud with no apparent motivation.
Secondly, the actual language itself contradicts
this possibility. While written in Aramaic and
Hebrew, there are only 3 words of likely Greek origins, and these are for
musical instruments. After what would be
160 years of Greek government rule, there should have been some invasion of the
Greek language in the text, as these intrusions, after so many years, generally
become a part of the language without regard to origin by the writer. However, there were at least 15 words of probably Persian
origin—3 of which were government positions that had apparently so long passed
from usage that when the transcript was later translated into Greek, that the
meanings could only be guessed at. These
terms were (roughly phonetically): dargazerayya (councilors), gedobrayya
(treasurers), and detabrayya (lawgivers)( Pictoral Encyclopedia of the Bible, vol 1, “Biblical
and Post-Biblical Aramaic” pg 254). If
indeed this prophetic book were a hoax that instead was accounting things that
had already passed, then how did it include such authentic language usage to
the point of identifying and eliminating all cultural and political words of
Greek origin? And how did a 2nd
century writer manage to research sufficiently to recreate the Persian language
influences to such detail that he used words no one knew the meaning of
anymore? Moreover, to what end?
Even this prophetic book, astonishingly
accurate, can be confirmed by outside evidence.
In reality, all the verifiable scientific evidence is consistent with
the Biblical accounts of the age of humanity, and how cultures spread from the
No other ancient text has recorded the extent of
world history that the Bible has, so much so that it is the main written
chronometer for ancient history. If all
the scientific evidence of these cultures irrefutably conflicted with the
Biblical account, then evolution would be the more rational model. But since this civilization timeline is only
authenticated by evolutionary assumptions, and not undeniable scientific certainty,
it must be rejected as a fact. There are
no facts in the archeological record that irrefutably conflict with the
timescale of the scientifically sound Creation/Flood model, and no skeletons to
support the ascent of man, just as there is not one fossil to document the
evolution of any life from lower forms.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE
INFO
Living Fossils
Evolutionists explain that we do not observe
evolution today because it happens gradually and painstakingly over millions of
years. Unfortunately, this explanation
is not supported by the fossil record.
Here is just a very short list of the hundreds of remarkable examples of
what evolutionists call “living fossils,” or species that have remained unchanged since
their supposedly ancient appearance in the fossil record, and still live
today:
Ants— today’s ants demonstrate a complete lack of
change from those found in amber from the Jurassic (contemporaneous with the
dinosaur)
Nautilus—the basic squid family has not “evolved” since
it first appeared, complete with eyes, in the Cambrian explosion—the first
level of evolution after single-celled organisms (along with numerous other
species still existing from that era, like starfish, and jellyfish, and clams,
and coral . . .)
Spider—appears complete 400 million years ago when
things were supposedly just getting out of the water. Wonder what they caught
in those fancy webs.
Dragonfly—and numerous plants and trees from the
carboniferous period, 100 million years before dinosaurs
Frog—275 million years ago
Cockroach—250 million years ago
Horseshoe Crab—Jurassic
Shark—400 million years ago
Tuatara—lizard-like reptiles that live in New
Zealand. They first appeared during the
dinosaur age, but no fossil of them has ever been found younger than “80
million years” –clearly the fossil record is unreliable for accurately recording
the span of an organism’s existence.
Coelacanth—appears in strata “200
million years” before dinosaurs, yet modern specimens show no evolution. Amazingly, according to evolutionists’ own
interpretation, this fish was deemed extinct for hundreds of millions years
based on their lack of appearance in the fossil record above that geologic
age. This is one of dozens of species
not traceable for millions of years of strata.
Clearly this animal succeeded throughout this supposedly tremendous
gap. Evolutionists view the fossil record completely through the
interpretive lens of evolution for advocating what and when things have lived,
no matter how unreliable this proves to be.
If such a distinct creature can live without leaving a trace
for hundreds of millions of years, then what is the record reliable for? If it can’t testify to the lifespan of a
species, how can it accurately record when species first appeared? Fossils, do tell a story, but
they do not tell the whole story.
Creationism completely explains such anomalies, since all the creatures
were only buried in one catastrophic flood event. The preserved remnant are but a
representation of the original diversity.
There are hundreds of examples of species, from bacteria to crocodiles (“140
million” years ago) that have not changed for supposedly millions of years from the
fossil record to the present. Despite
its abundant confirmation of stasis, the fossil record cannot offer any
evidence of evolutionary development. It
is as if each species came into being in an instant, fully formed. In reality, when we look at all the thousands
of species of plants, animals, arthropods, and more, we must acknowledge that
all these unique species would have been in a constant state of change in order
to make these millions of micro-steps in all their different directions.
It is simply not biologically possible, or even
logical, that the past was a state of constant unending flow of transitions
which miraculously resulted not only in the species we still have, but the
species already lost to extinction. It
is unimaginable that all this happened for hundreds of millions of years in all
these thousands of species, and not one true transition is
recorded in the fossil record. How is it
that the supposed turbulent past of constant change that we cannot
witness, does not in any way resemble the present that we can observe. Moreover, the record itself provides
evidence, species after species, of stability for “hundreds of millions of
years,” up until the present.
The true facts, as presented by the fossil
record, validate the Creation model to the complete exclusion of the theory of
evolution. It demonstrates that the
“Cambrian System” was instantly crowded with fully developed and familiar
species when they first appear. Even though
this is the first opportunity for evolution to show off, there are no fossils
to demonstrate how dozens of new body types emerged from the single-celled
organisms.
The change from invertebrates to vertebrates is
just as mysterious. The fossils manage
to clearly record the instantaneous wide variety of fish, but no developmental
stages. The fossil record continues to
point to stasis, and does not offer evidence of how a fish turned into such a
variety of amphibians, straining our imaginations. All the way through the “rise” of mammals,
birds, and humans, the abundant fossils faithfully preserve only the terminal,
or final species, omitting the millions of transitional species.
All species are found utterly complete in every
way when they are uncovered from the sand and mud and limestone that entombed
them. We will never know how evolution
could haphazardly produce such marvels as an eye, or wings, or flowers, or any
complex feature because their fossils just appear again with no recorded
developmental history in spite of all the suitable layers of sediments. All the uniqueness of life, from birds, to
marsupials, and hundreds of unusual specialized traits, are captured in these
rocks as if they just materialized, already fashioned, without any ancestry. In fact, a much greater variety, even giant
forms, of species evidently lived in the past, attesting to the dominant value
of extinction and entropy, not evolution.
Creationists are not surprised that the fossil
record testifies against the gradual transitions of evolution, because
Special Creation would leave a record of stasis in all species because
they were created at the same time.
These were the forms buried in one sudden cataclysm as a testimony of
this flood, and left for our examination.
Clearly the world is winding down, not
elaborating. According to the fossil
record, there are less species now than ever, and species have not grown more
complex. Nothing in the fossil record
substantiates evolution. That scientists
continue to imply that there is solid evidence in the rocks is deceptive to
both the public, and themselves. The
fossil record offers no more evidentiary basis for evolution than a cemetery
does. In fact, evolution has no more
basis in reality than that Zeus sits atop
Ronald R. West wrote, and many evolutionists
concur, in the May, 1968 edition of Compass:
Contrary
to what most scientists write, the fossil record does not support the Darwinian
theory of evolution because it is this theory which we use to interpret the
fossil record. By doing so, we are
guilty of circular reasoning if we then say the fossil record supports this
theory.
Scientists see evolution in the fossils because
the fossils are essential to evolution.
INTRO PHYSICS BIOLOGY back to top GEOLOGY CHALLENGE INFO